生物膜的组成与结构
图片预览





文档简介
(共36张PPT)
第一节 生物膜的组成和结构
一、 概念
生物膜是构成细胞的膜系统的总称,包括围在细胞质外围的质膜和细胞器的内膜系统。
膜结构使细胞的生命活动与代谢过程有序进行
许多生理/病理过程,涉及到膜结构/功能的变化
外周膜
内膜系统
二、 化 学 组 成
膜 脂
膜 蛋 白
糖 类
无 机 盐
金 属 离 子
水
1. 膜 脂
磷 脂(主要)
糖 脂
甾 醇
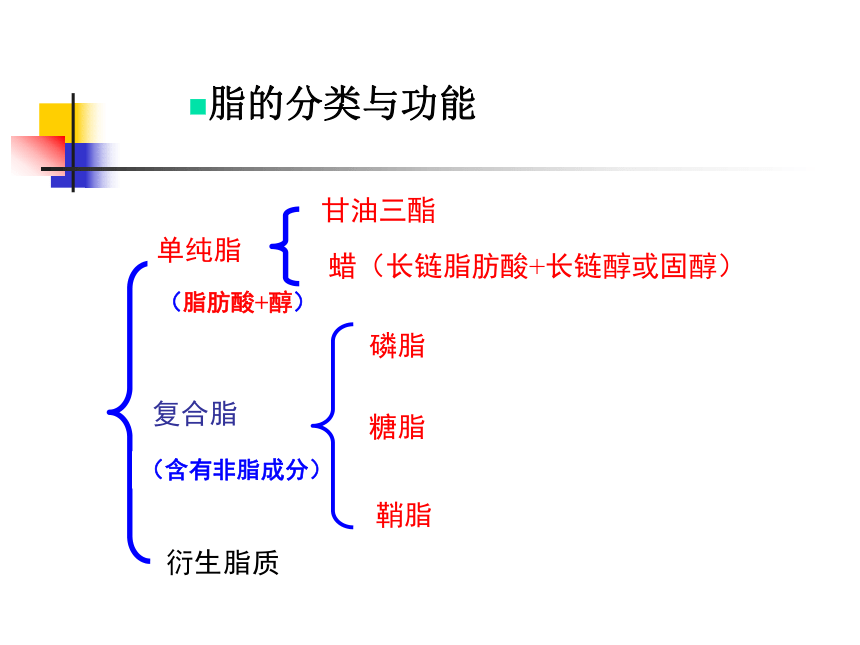
脂的分类与功能
单纯脂
复合脂
(含有非脂成分)
(脂肪酸+醇)
甘油三酯
衍生脂质
蜡(长链脂肪酸+长链醇或固醇)
磷脂
糖脂
鞘脂
2.体表脂类保护作用;
提问:脂类有哪些功能?
1.能量物质
三酰甘油脂又称油脂,每克的发热值比同质量的糖、蛋白质高2.3倍,并且不溶于水,在细胞内易于聚集,储存,故而被普遍作为细胞的能量储备物质。
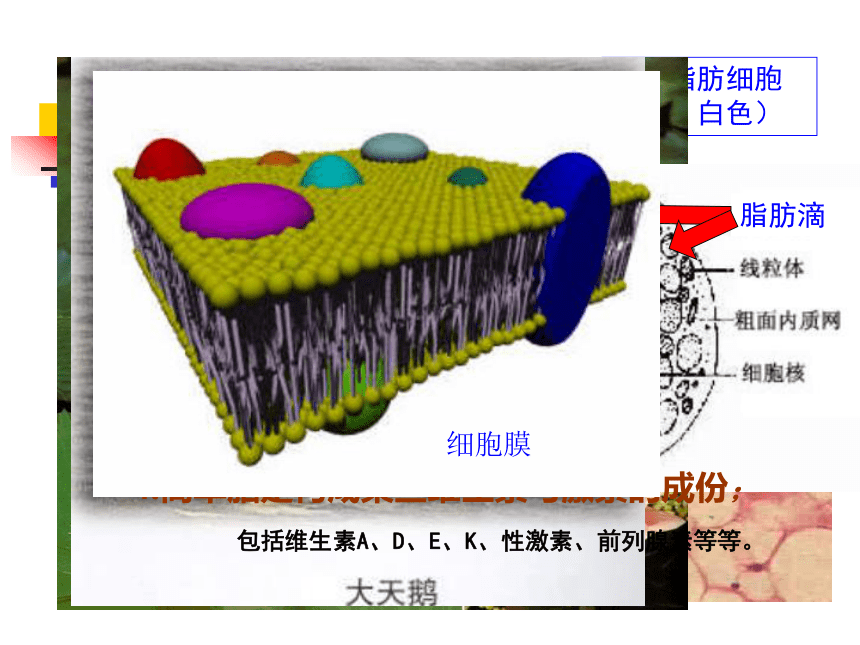
皮下脂肪细胞(黄、白色)
细胞核
细胞质
脂肪滴
脂肪滴
3.脂类是细胞膜的主要成份;
4.简单脂是构成某些维生素与激素的成份;
包括维生素A、D、E、K、性激素、前列腺素等等。
细胞膜
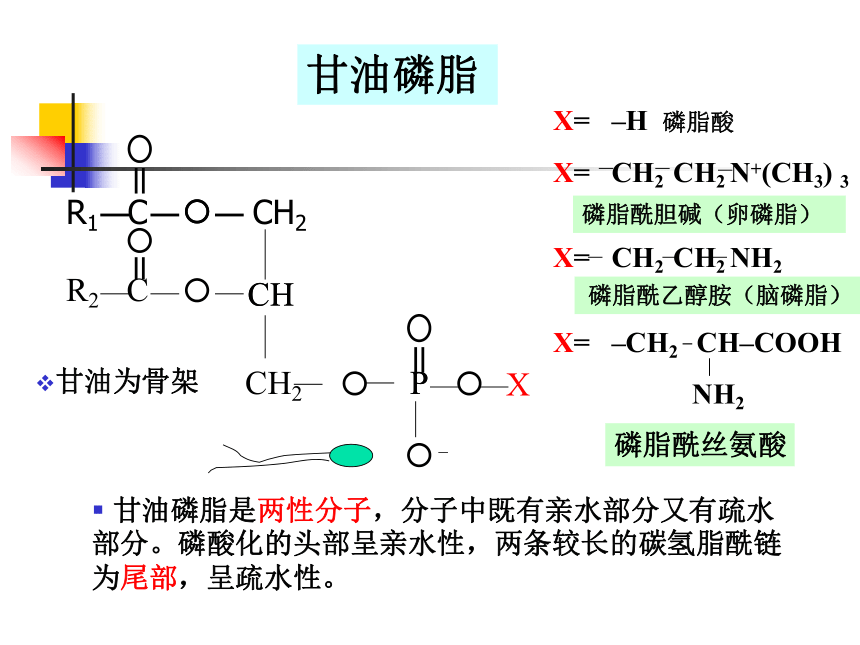
⑴ 磷 脂
甘油磷脂(主要)
鞘磷脂
R1 C CH2
CH
R2 C
CH
P
CH2
P
_
X
X= –H 磷脂酸
X= CH2 CH2 N+(CH3) 3
X= CH2 CH2 NH2
X= –CH2 CH–COOH
NH2
磷脂酰胆碱(卵磷脂)
磷脂酰乙醇胺(脑磷脂)
甘油磷脂
甘油磷脂是两性分子,分子中既有亲水部分又有疏水部分。磷酸化的头部呈亲水性,两条较长的碳氢脂酰链为尾部,呈疏水性。
磷脂酰丝氨酸
甘油为骨架
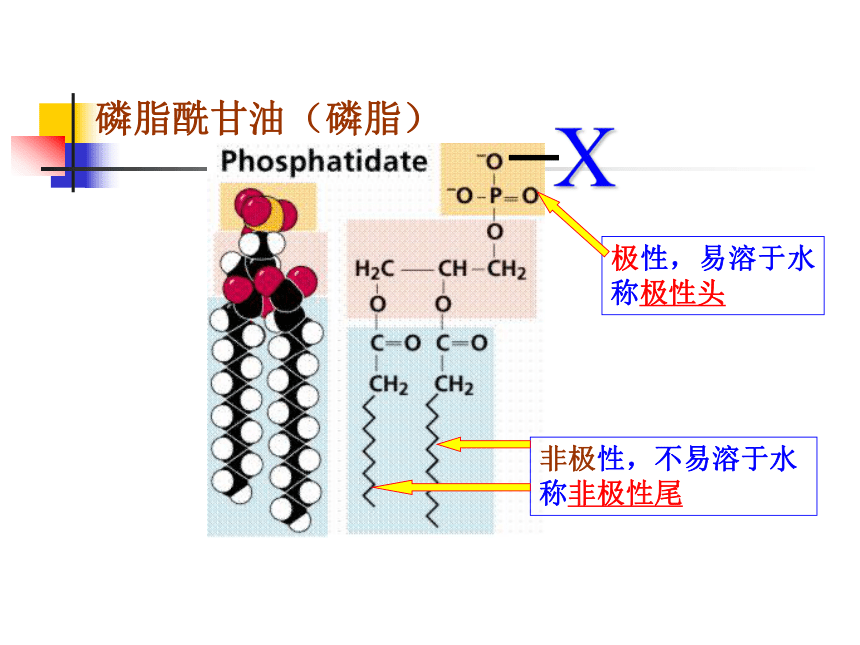
磷脂酰甘油(磷脂)
X
非极性,不易溶于水
称非极性尾
极性,易溶于水称极性头
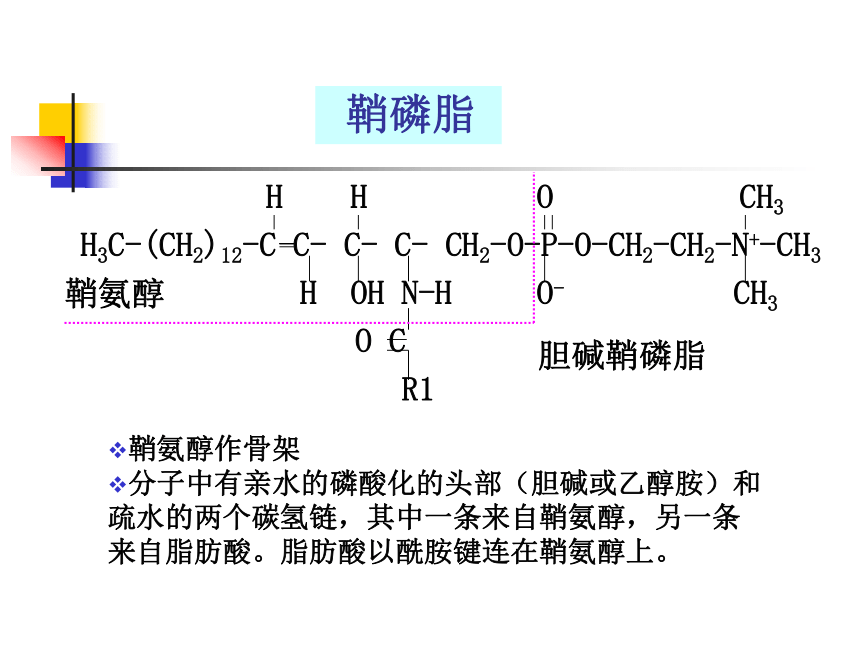
鞘磷脂
H H O CH3
H3C-(CH2)12-C C- C- C- CH2-O-P-O-CH2-CH2-N+-CH3
H OH N-H O- CH3
O C
R1
鞘氨醇
鞘氨醇作骨架
分子中有亲水的磷酸化的头部(胆碱或乙醇胺)和疏水的两个碳氢链,其中一条来自鞘氨醇,另一条来自脂肪酸。脂肪酸以酰胺键连在鞘氨醇上。
胆碱鞘磷脂
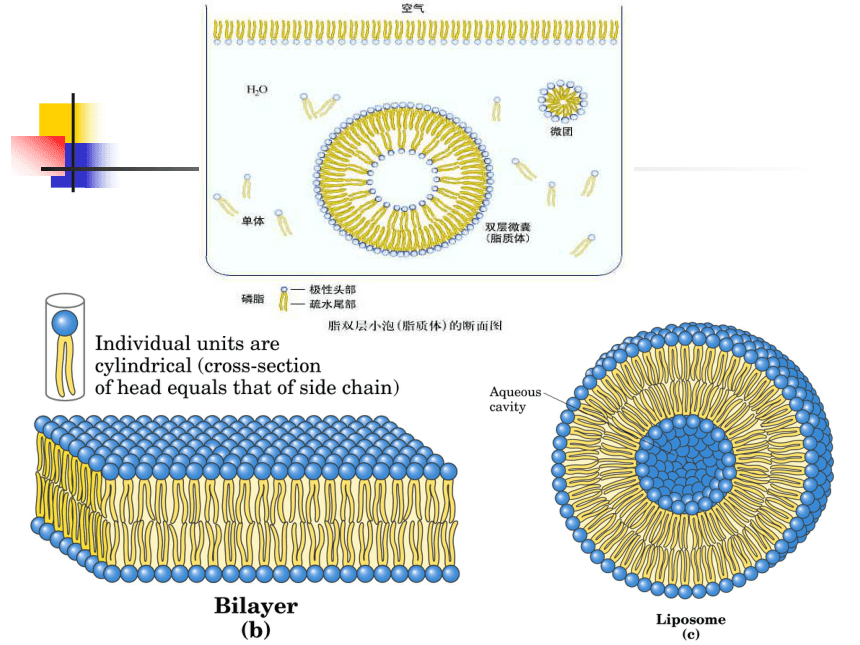
由磷脂形成的双层脂膜的示意图
在水溶液中两性的磷脂分子为避免疏水部分接触水分子而定向排列,形成脂双层结构。脂双层中,磷脂分子的疏水基团在内部而亲水的头部在表面。
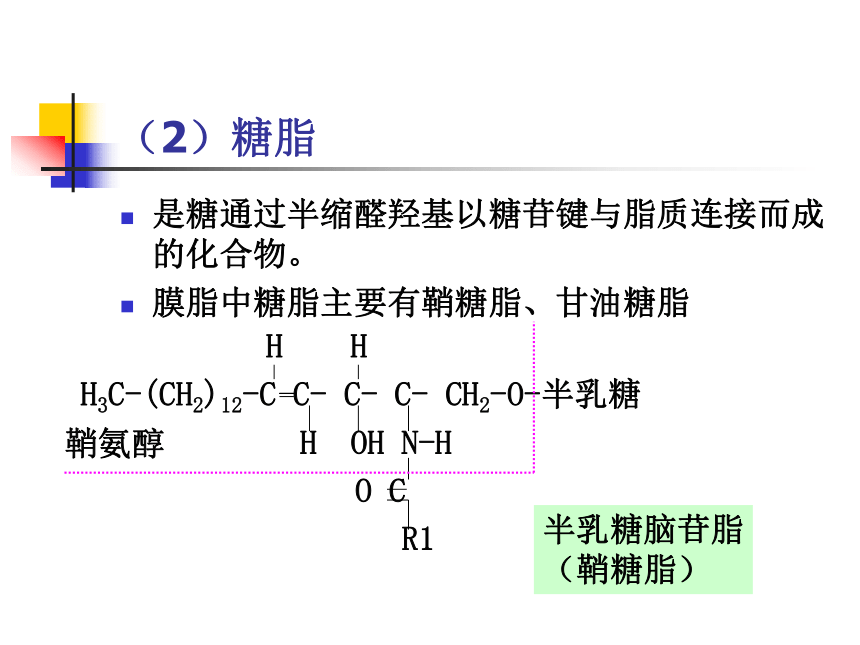
(2)糖脂
是糖通过半缩醛羟基以糖苷键与脂质连接而成的化合物。
膜脂中糖脂主要有鞘糖脂、甘油糖脂
H H
H3C-(CH2)12-C C- C- C- CH2-O-半乳糖
H OH N-H
O C
R1
鞘氨醇
半乳糖脑苷脂
(鞘糖脂)
(3) 甾 醇
又名固醇,动物膜甾醇主要是胆固醇,植物膜甾醇含量较动物少,主要有谷甾醇、豆甾醇等。
2. 膜蛋白
⑴内 在 蛋 白
⑵外 周 蛋 白
有的全部埋于脂双层的疏水区,有的部分嵌在脂双层中,有的横跨全膜.主要靠疏水作用通过某些非极性氨基酸残基与膜脂疏水部分相结合。
这类蛋白被紧密连在膜上,并且不易溶于水。只有用破坏膜结构的试剂如有机溶剂(氯仿)或去污剂(TritonX-100)才能把它们从膜中提取出来。
(1)内在蛋白
膜锚蛋白
有些膜内在蛋白本身并没有进入膜内,他们以共价键与脂质、脂酰链、异戊烯基团相结合并通过他们的疏水部分插入到膜内,这种形式的内在蛋白称为膜锚蛋白。
(2) 外 周 蛋 白
分布于膜的脂双层内外表面,通过极性氨基酸残基以离子键、氢键、范德华力等次级键与膜脂极性头部或与内在蛋白的亲水部分结合。
比较易于分离,大都能溶于水,可在不破坏膜结构的情况下,通过温和方法(高离子强度、高pH、)分离提取。
外周蛋白和膜内 (嵌入)蛋白
外周蛋白
嵌入(膜内)蛋白
去污剂
糖蛋白
pH改变、螯合剂、尿素、碳酸盐可除去外周蛋白
3. 糖类
生物膜中的糖类大多与膜蛋白结合 糖蛋白(信息识别)
少数与膜脂结合 糖脂
糖类在膜上的分布
非对称的,全部分布在膜的非细胞质一侧。
质膜上的糖
细胞内膜的糖
糖蛋白,糖脂
三 生物膜的分子结构模型
流体镶嵌模型
1972年美国Singer和Nicolson提出,认为生物膜是一种流动的、嵌有各种蛋白质的脂质双分子层结构,其中蛋白质犹如一座座冰山漂移在流动脂质的海洋中。
与过去模型的主要差别
突出了膜的流动性
显示了膜蛋白分布的不对称性
四、 生 物 膜 的 特 性
㈠ 膜分子结构的不对称性
㈡ 膜分子结构的流动性
㈠ 膜分子结构的不对称性
1、膜脂的分布不对称,即膜脂双分子层内外两侧的脂种类、含量不同,如人红细胞质膜:
膜的外层卵磷脂、鞘磷脂较多
膜的内层脑磷脂、磷脂酰丝氨酸较多
2、膜蛋白的分布不对称
如线粒体内膜中的NADH电子传递链各组分:
Cyt氧化酶 、琥珀酸脱氢酶在线粒体内膜内侧
Cytc在线粒体内膜外侧
3、糖蛋白和糖脂中的多糖只分布在
膜的非细胞质一侧
线粒体
内
外
(二)膜分子结构的流动性
膜的流动性主要是指膜脂及膜蛋白流动性。
合适的流动性对生物膜表现其正常功能十分重要.
膜脂的流动性
膜脂的流动性主要决定于磷脂分子.
在生理条件下,磷脂大多呈流动的液晶态,磷脂在膜内可作旋转运动,翻转运动,侧向运动等.当温度降至一定值时,膜脂从流动的液晶态转变为类似晶体的凝胶态,这个温度称为相变温度.凝胶状态也可再熔解为液晶态。各种膜脂由于组分不同而具有各自的相变温度。
温度引起侧链热运动
脂双层平面的扩散
跨膜扩散:“翻跟头”
膜脂的运动
膜脂的流动性的大小与磷脂分子中脂肪酸链的长短及不饱和程度密切相关.链越短,不饱和程度越高,流动性越大.
哺乳动物中胆固醇对膜脂流动性也有一定的调控作用,在生理条件下增加胆固醇的含量会降低膜的流动性,因为胆固醇的闭合环状结构干扰了脂肪酸的侧向运动。
膜脂的流动性是不均匀的,在一定温度下,有的膜脂处于凝胶态,有的则呈液晶态,处于液晶态的各膜脂的流动性也不完全相同.
膜蛋白的流动性
膜蛋白能做侧向扩散和旋转扩散,其速度平均比膜脂小10-100倍.
膜蛋白的扩散运动
膜的融合
生物膜的一个明显的特征是可与另一个膜融合而不失去其完整性。膜虽是稳定的,但不是静止的,内膜系统中膜状结构不断地从高尔基复合体上分泌形成;外吞、内饮、细胞分裂、精卵细胞融合、膜包裹病毒进入宿主细胞等都涉及膜的重新形成,而它们最基本的行为就是两个膜片段的融合而不失去完整性。
膜的融合(Membrrane Fusion)
第一节 生物膜的组成和结构
一、 概念
生物膜是构成细胞的膜系统的总称,包括围在细胞质外围的质膜和细胞器的内膜系统。
膜结构使细胞的生命活动与代谢过程有序进行
许多生理/病理过程,涉及到膜结构/功能的变化
外周膜
内膜系统
二、 化 学 组 成
膜 脂
膜 蛋 白
糖 类
无 机 盐
金 属 离 子
水
1. 膜 脂
磷 脂(主要)
糖 脂
甾 醇
脂的分类与功能
单纯脂
复合脂
(含有非脂成分)
(脂肪酸+醇)
甘油三酯
衍生脂质
蜡(长链脂肪酸+长链醇或固醇)
磷脂
糖脂
鞘脂
2.体表脂类保护作用;
提问:脂类有哪些功能?
1.能量物质
三酰甘油脂又称油脂,每克的发热值比同质量的糖、蛋白质高2.3倍,并且不溶于水,在细胞内易于聚集,储存,故而被普遍作为细胞的能量储备物质。
皮下脂肪细胞(黄、白色)
细胞核
细胞质
脂肪滴
脂肪滴
3.脂类是细胞膜的主要成份;
4.简单脂是构成某些维生素与激素的成份;
包括维生素A、D、E、K、性激素、前列腺素等等。
细胞膜
⑴ 磷 脂
甘油磷脂(主要)
鞘磷脂
R1 C CH2
CH
R2 C
CH
P
CH2
P
_
X
X= –H 磷脂酸
X= CH2 CH2 N+(CH3) 3
X= CH2 CH2 NH2
X= –CH2 CH–COOH
NH2
磷脂酰胆碱(卵磷脂)
磷脂酰乙醇胺(脑磷脂)
甘油磷脂
甘油磷脂是两性分子,分子中既有亲水部分又有疏水部分。磷酸化的头部呈亲水性,两条较长的碳氢脂酰链为尾部,呈疏水性。
磷脂酰丝氨酸
甘油为骨架
磷脂酰甘油(磷脂)
X
非极性,不易溶于水
称非极性尾
极性,易溶于水称极性头
鞘磷脂
H H O CH3
H3C-(CH2)12-C C- C- C- CH2-O-P-O-CH2-CH2-N+-CH3
H OH N-H O- CH3
O C
R1
鞘氨醇
鞘氨醇作骨架
分子中有亲水的磷酸化的头部(胆碱或乙醇胺)和疏水的两个碳氢链,其中一条来自鞘氨醇,另一条来自脂肪酸。脂肪酸以酰胺键连在鞘氨醇上。
胆碱鞘磷脂
由磷脂形成的双层脂膜的示意图
在水溶液中两性的磷脂分子为避免疏水部分接触水分子而定向排列,形成脂双层结构。脂双层中,磷脂分子的疏水基团在内部而亲水的头部在表面。
(2)糖脂
是糖通过半缩醛羟基以糖苷键与脂质连接而成的化合物。
膜脂中糖脂主要有鞘糖脂、甘油糖脂
H H
H3C-(CH2)12-C C- C- C- CH2-O-半乳糖
H OH N-H
O C
R1
鞘氨醇
半乳糖脑苷脂
(鞘糖脂)
(3) 甾 醇
又名固醇,动物膜甾醇主要是胆固醇,植物膜甾醇含量较动物少,主要有谷甾醇、豆甾醇等。
2. 膜蛋白
⑴内 在 蛋 白
⑵外 周 蛋 白
有的全部埋于脂双层的疏水区,有的部分嵌在脂双层中,有的横跨全膜.主要靠疏水作用通过某些非极性氨基酸残基与膜脂疏水部分相结合。
这类蛋白被紧密连在膜上,并且不易溶于水。只有用破坏膜结构的试剂如有机溶剂(氯仿)或去污剂(TritonX-100)才能把它们从膜中提取出来。
(1)内在蛋白
膜锚蛋白
有些膜内在蛋白本身并没有进入膜内,他们以共价键与脂质、脂酰链、异戊烯基团相结合并通过他们的疏水部分插入到膜内,这种形式的内在蛋白称为膜锚蛋白。
(2) 外 周 蛋 白
分布于膜的脂双层内外表面,通过极性氨基酸残基以离子键、氢键、范德华力等次级键与膜脂极性头部或与内在蛋白的亲水部分结合。
比较易于分离,大都能溶于水,可在不破坏膜结构的情况下,通过温和方法(高离子强度、高pH、)分离提取。
外周蛋白和膜内 (嵌入)蛋白
外周蛋白
嵌入(膜内)蛋白
去污剂
糖蛋白
pH改变、螯合剂、尿素、碳酸盐可除去外周蛋白
3. 糖类
生物膜中的糖类大多与膜蛋白结合 糖蛋白(信息识别)
少数与膜脂结合 糖脂
糖类在膜上的分布
非对称的,全部分布在膜的非细胞质一侧。
质膜上的糖
细胞内膜的糖
糖蛋白,糖脂
三 生物膜的分子结构模型
流体镶嵌模型
1972年美国Singer和Nicolson提出,认为生物膜是一种流动的、嵌有各种蛋白质的脂质双分子层结构,其中蛋白质犹如一座座冰山漂移在流动脂质的海洋中。
与过去模型的主要差别
突出了膜的流动性
显示了膜蛋白分布的不对称性
四、 生 物 膜 的 特 性
㈠ 膜分子结构的不对称性
㈡ 膜分子结构的流动性
㈠ 膜分子结构的不对称性
1、膜脂的分布不对称,即膜脂双分子层内外两侧的脂种类、含量不同,如人红细胞质膜:
膜的外层卵磷脂、鞘磷脂较多
膜的内层脑磷脂、磷脂酰丝氨酸较多
2、膜蛋白的分布不对称
如线粒体内膜中的NADH电子传递链各组分:
Cyt氧化酶 、琥珀酸脱氢酶在线粒体内膜内侧
Cytc在线粒体内膜外侧
3、糖蛋白和糖脂中的多糖只分布在
膜的非细胞质一侧
线粒体
内
外
(二)膜分子结构的流动性
膜的流动性主要是指膜脂及膜蛋白流动性。
合适的流动性对生物膜表现其正常功能十分重要.
膜脂的流动性
膜脂的流动性主要决定于磷脂分子.
在生理条件下,磷脂大多呈流动的液晶态,磷脂在膜内可作旋转运动,翻转运动,侧向运动等.当温度降至一定值时,膜脂从流动的液晶态转变为类似晶体的凝胶态,这个温度称为相变温度.凝胶状态也可再熔解为液晶态。各种膜脂由于组分不同而具有各自的相变温度。
温度引起侧链热运动
脂双层平面的扩散
跨膜扩散:“翻跟头”
膜脂的运动
膜脂的流动性的大小与磷脂分子中脂肪酸链的长短及不饱和程度密切相关.链越短,不饱和程度越高,流动性越大.
哺乳动物中胆固醇对膜脂流动性也有一定的调控作用,在生理条件下增加胆固醇的含量会降低膜的流动性,因为胆固醇的闭合环状结构干扰了脂肪酸的侧向运动。
膜脂的流动性是不均匀的,在一定温度下,有的膜脂处于凝胶态,有的则呈液晶态,处于液晶态的各膜脂的流动性也不完全相同.
膜蛋白的流动性
膜蛋白能做侧向扩散和旋转扩散,其速度平均比膜脂小10-100倍.
膜蛋白的扩散运动
膜的融合
生物膜的一个明显的特征是可与另一个膜融合而不失去其完整性。膜虽是稳定的,但不是静止的,内膜系统中膜状结构不断地从高尔基复合体上分泌形成;外吞、内饮、细胞分裂、精卵细胞融合、膜包裹病毒进入宿主细胞等都涉及膜的重新形成,而它们最基本的行为就是两个膜片段的融合而不失去完整性。
膜的融合(Membrrane Fusion)
同课章节目录
- 第一章 走近细胞
- 第1节 从生物圈到细胞
- 第2节 细胞的多样性和统一性
- 第二章 组成细胞的分子
- 第1节 细胞中的元素和化合物
- 第2节 生命活动的主要承担者──蛋白质
- 第3节 遗传信息的携带者──核酸
- 第4节 细胞中的糖类和脂质
- 第5节 细胞中的无机物
- 第三章 细胞的基本结构
- 第1节 细胞膜──系统的边界
- 第2节 细胞器──系统内的分工合作
- 第3节 细胞核──系统的控制中心
- 第四章 细胞的物质输入和输出
- 第1节 物质跨膜运输的实例
- 第2节 生物膜的流动镶嵌模型
- 第3节 物质跨膜运输的方式
- 第五章 细胞的能量供应和利用
- 第1节 降低化学反应活化能的酶
- 第2节 细胞的能量“通货”──ATP
- 第3节 ATP的主要来源──细胞呼吸
- 第4节 能量之源——光与光合作用
- 第六章 细胞的生命历程
- 第1节 细胞的增殖
- 第2节 细胞的分化
- 第3节 细胞的衰老和凋亡
- 第4节 细胞的癌变