浙科版必修3第三章第二节特异性免疫反应(共43张PPT)
文档属性
| 名称 | 浙科版必修3第三章第二节特异性免疫反应(共43张PPT) |

|
|
| 格式 | zip | ||
| 文件大小 | 1.8MB | ||
| 资源类型 | 教案 | ||
| 版本资源 | 浙科版 | ||
| 科目 | 生物学 | ||
| 更新时间 | 2014-09-09 00:00:00 | ||
图片预览







文档简介
课件43张PPT。小测验1、构成非特异性免疫的因素有哪些?
2、什么是补体?
3、补体的经典途径如何?
4、补体激活后具有的生物学功能?第四章 特异性免疫
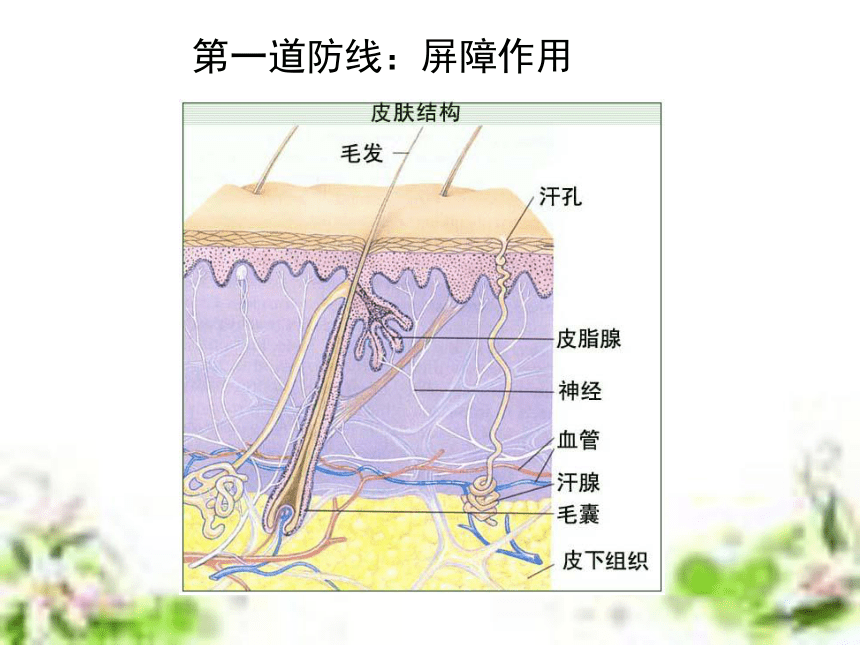
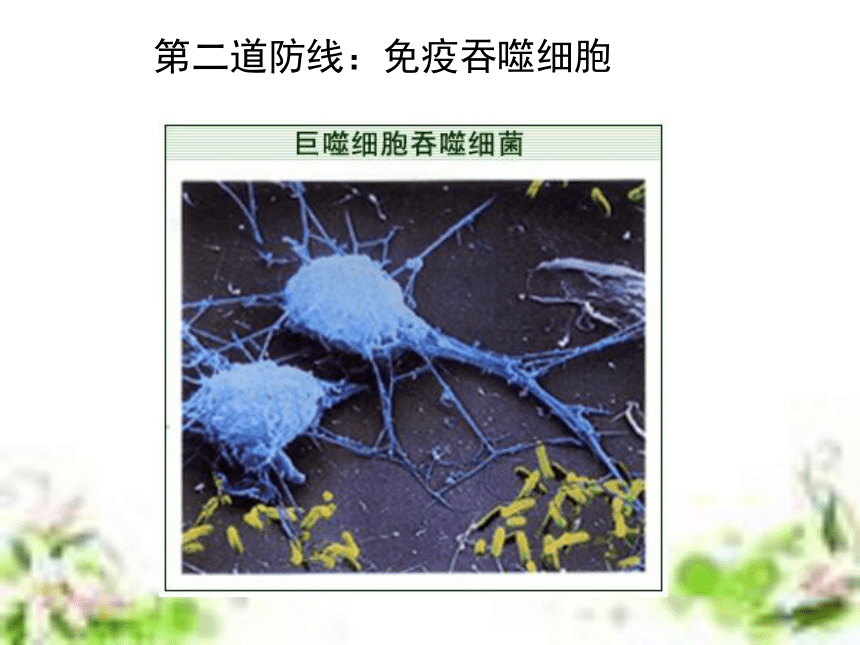
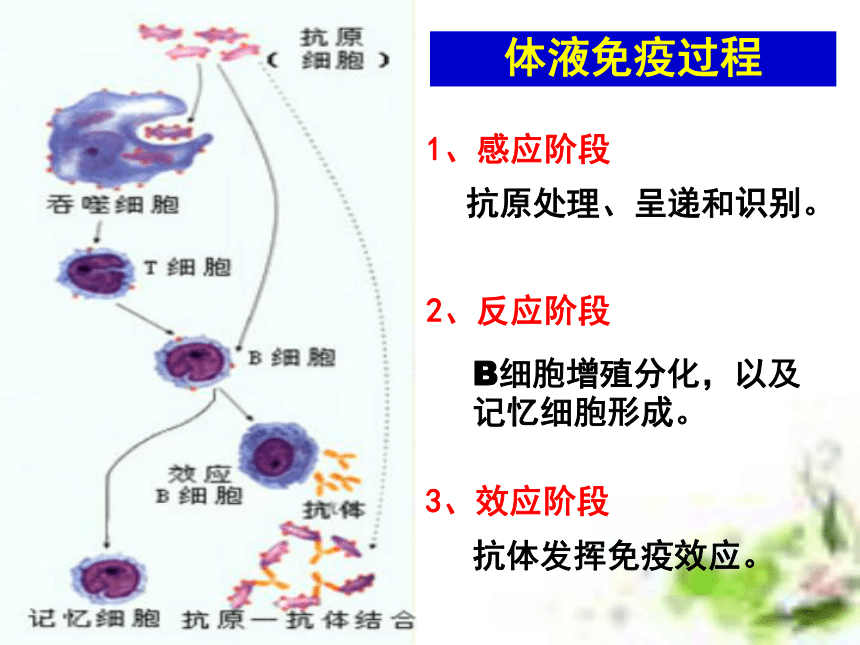
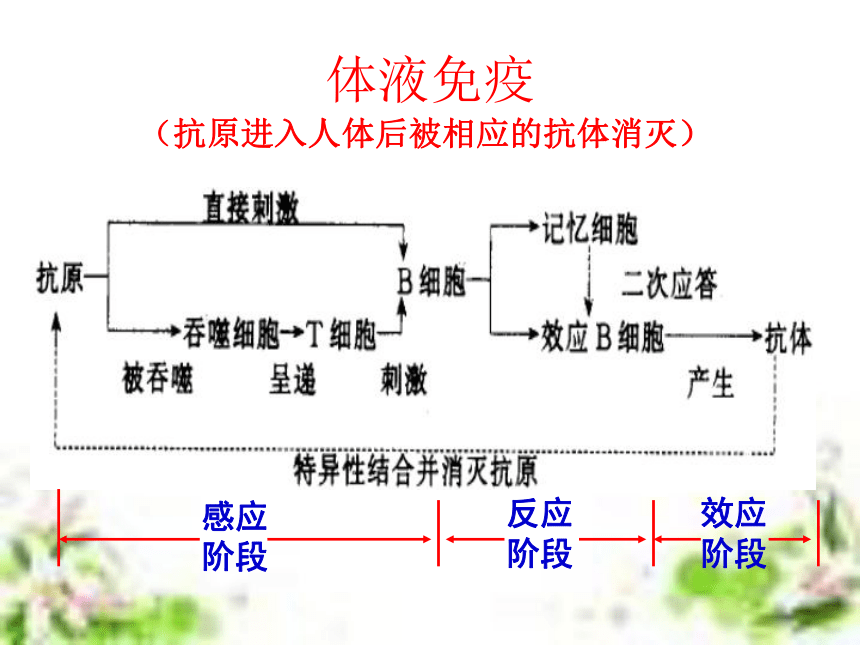
(6学时) 非典型肺炎流行猖獗引起社会的高度重视!但即使是流行最严重的地区,也不是人人都会得病!因为,冠状病毒虽然厉害,对于免疫功能强大的人,它也是无能为力的!目前全世界有大约6000万人感染了艾滋病毒,但从80年代中期以来,科学家发现,约有2%的病毒携带者竟然终身没有发展成艾滋病,成为“长期存活的病毒携带者”。这一现像表明,部份人对艾滋病毒具有先天的免疫力。津巴布韦一名6岁的艾滋病患儿第一道防线:屏障作用 人体的皮肤和黏膜等组成人体的第一道防线第二道防线:免疫吞噬细胞第三道防线:补体和溶菌酶等 特异性免疫应答是动物在个体发育过程中,由于受抗原异物的刺激,而发展生成有针对性的免疫力,包括引起生成致敏淋巴细胞的特异性细胞免疫和产生抗体所表现出的特异性体液免疫。这种免疫应答生成的原因,是由于机体与入侵的病原微生物或其他各种抗原异物相互斗争,或因接种疫苗,或因无意接触外界抗原异物后,引起T细胞及B细胞发生免疫应答而生成的。 特异性免疫的基本概念特异性免疫应答与非特异性免疫应答比较,它具有三个主要特点:①不能遗传给后代,它是在个体发育中,在非特异性免疫的基础上发展生成的,故属于后天获得性免疫。②具有严格的特异性,即它只对于引起其产生这种免疫力的同种抗原异物有对抗性,对于无关的抗原物质不起作用,故属于特异性免疫。③特异性免疫应答是在非特异性免疫应答的基础上建立起来的,也就是说,它需要非特异的体液因子及细胞因子相互配合协作,才能发挥更强大的免疫效能,既可表现免疫防护效果,又可引起免疫病理损害。 特异性免疫的基本过程 在抗原刺激下,机体形成特异性免疫应答,不论细胞免疫或体液免疫,都有其共同的规律,一般分为感应(识别)即加工处理抗原阶段、反应即淋巴细胞分化增殖阶段和效应即发生免疫应答(产生抗体或致敏淋巴细胞再次遇到抗原后刺激产生淋巴因子)三个阶段。1、感应阶段2、反应阶段3、效应阶段体液免疫过程抗原处理、呈递和识别。B细胞增殖分化,以及记忆细胞形成。抗体发挥免疫效应。体液免疫(抗原进入人体后被相应的抗体消灭)细胞免疫过程1、感应阶段2、反应阶段3、效应阶段抗原处理、呈递和识别。T细胞增殖分化,以及记忆细胞形成。效应T细胞、抗体和淋巴因子发挥免疫效应。细胞免疫(抗原进入人体后被相应的免疫细胞消灭) 淋巴细胞系(T细胞、B细胞、效应T细胞、效应B细胞)
吞噬细胞系(巨噬细胞,肥大细胞)。
粒细胞系(噬中性粒细胞、噬酸性粒细胞、噬碱性粒细胞)免疫细胞淋巴细胞的分化 实验探索强X光照射去 胸腺小鼠无免疫性输给小鼠来自胸腺的淋巴细胞 免疫功能得到恢复,但不能产生游离于体液中的抗体 输给来自骨髓的淋巴细胞 免疫功能不能恢复,但产生抗体的功能得到一定恢复 同时输入来自胸腺和骨髓的淋巴细胞小鼠就可恢复
全部免疫功能 实验结论: 胸腺内的淋巴细胞和骨髓中的淋巴细胞在免疫中起不同的作用,分别称为T细胞和B细胞。体液免疫细胞免疫体液免疫与细胞免疫的关系 两者各自有其独特作用,又可相互协作,共同发挥作用。各阶段注意点1、感应阶段: 吞噬细胞的作用是使抗原的抗原决定簇暴露,各细胞间的呈递是通过细胞的相互接触来完成的。2、反应阶段: 形成的记忆细胞在同种抗原再次侵入时,可迅速增殖、分化,形成大量的效应细胞,产生更强的特异性免疫反应。①体液免疫中,抗体与病菌结合后,可抑制其繁殖和对宿主细胞的黏附,防止感染和疾病的发生。抗体与病毒结合后,可使其失去侵染和破坏宿主细胞的能力。抗原和抗体结合后进一步的变化是形成沉淀或细胞集团,进而被吞噬细胞吞噬消化。3、效应阶段:②细胞免疫中,效应T细胞与被抗原入侵的宿主细胞(即靶细胞)接触,激活靶细胞的溶酶体酶,最终使靶细胞裂解死亡。细胞内的抗原也因失去藏身之所而为抗体消灭。 抗体抗体发现的过程:
19世纪末20世纪初,科学家们在实验中发现,用细菌或其外毒素给动物注射,过一段时间后,该动物的血清中出现一些有防御作用的保护性成分。
在20世纪30年代,科学家用电泳法鉴定,证明抗体是一种球蛋白。
1959年科学家对抗体的结构进行了研究,证明它是由四条肽链组成,借二硫键连接在一起,形成一个"Y"型的四链分子。不同的抗体某些部分氨基酸序列各不相同,决定抗体的特异性。 具有抗体活性的动物蛋白。主要存在于血浆中,也见于其他体液、组织和一些分泌液中。人血浆内的免疫球蛋白大多数存在于丙种球蛋白(γ-球蛋白)中。免疫球蛋白(Ig)Ig分子根据其可溶性、电荷、大小、化学结构等理化特性及其免疫反应性可分为IgA,IgG,IgD,IgE和IgM五类。在电镜下,IgG为“Y”型分子。“Y”的臂可结合抗原。若用化学方法处理免疫球蛋白,使其摆脱二硫键的束缚,分子就被裂解成4条独立的多肽链。所有Ig都具有由四条肽链组成的基本结构,称为单体(抗体单位),IgG为典型的单体。 抗体(antibody,Ab)是介导体液免疫的重要效应分子,是B细胞接受抗原刺激后增殖分化为浆细胞所产生的一种糖蛋白,主要存在于血清等体液中,能与相应抗原特异地结合,具有免疫功能。 抗体的概念 Porter对血清IgG抗体的研究证明,Ig分子基本结构是由四个肽链组成的,二条较小的轻链和二条较大的重链,轻链与重链是由二硫键连接形成,分为氨基端(N端),羧基端(C端)。任何一类天然免疫球蛋白分子均含有四条异源性多肽链,其中,相对分子质量较大的称为重链(heavy chain,H链),而相对分子质量较小的为轻链(light chain,L链)。同一天然Ig分子中的两条H链和两条L链的氨基酸组成完全相同。 重链和轻链重链:两条相同的长链称为重链、简称H链。H链由450—550氨基酸组成,相对分子质量为50—75kDa。根据Ig重链恒定区氨基酸的排列顺序不同,免疫反应性就不同,据此可将血清中的Ig分成五大类:IgG,IgA,IgM,IgD,IgE,其相应的重链分别为γ、μ、α、δ和ε链。 轻链:两条相同的短链称为轻链,简称L链,L链约由214个氨基酸组成,相对分子质量约为25kDa,根据L链免疫反应性不同,L链可分为两型,即κ型和λ型。根据λ链恒定区个别氨基酸的差异,又可分为λ1、λ2、λ3和λ4四个亚基(subtype)。同一天然的Ig分子上两条H链是同类,两条轻链是同型,五类Ig中,每一类Ig都可以有κ链或λ链,可以把Ig分成五类十型。 重链和轻链 可变区和恒定区 免疫球蛋白轻链和重链中靠近N端氨基酸序列变化较大的区域称为可变区(variable region,V区),分别占重链和轻链的1/4和1/2;而靠近C端氨基酸序列相对稳定的区域,称为恒定区(constant region,C区),分别占重链和轻链的3/4和1/2。可变区:免疫球蛋白轻链靠近氨基端(N端)的1/2区段或重链靠近氨基端(N端)1/5—1/4区段的110个氨基酸的组成和排列顺序在不同Ig中差异很大,称为可变区,分别用VH和VL表示。 可变区和恒定区 高变区(hypervariable region,HVR):在VH和VL中,各有3个区域的氨基酸组成和排列顺序高度可变,称为高变区或互补决定区(complementarity determining region,CDR),分别用HVRl(CDRl),HVR2(CDR2),HVR3(CDR3)表示。 恒定区:免疫球蛋白除了可变区以外,轻链靠近羧基端(C端)1/2区段或重链靠近C端3/4—4/5区段氨基酸的组成和排列相对稳定,称为恒定区。分别用CH和CL表示,不同Ig的CL的长度基本一致;不同Ig的CH长度不一,有的包括CH1,CH2,CH3或CH1,CH2,CH3,CH4。铰链区:位于CH1和CH2之间的非独立功能区,即位于Ig的Y形分子两臂和主干之间的区域,称为铰链区。富含脯氨酸,因此易伸展弯曲,易被木瓜蛋白酶、胃蛋白酶水解,并可通过变构有利于与不同距离的抗原决定簇结合。 可变区和恒定区铰链区的作用:
(1)当VL,VH与抗原结合时,此区发生扭曲,使抗体分子上两个抗原结合位点更好地与两个抗原决定簇发生互补,利于抗体分子高变区与抗原决定簇吻合。
(2)Ag和Ab结合后,铰链区发生构型变化,使存在于CH2区补体结合位点暴露,使补体得以结合,显示出活化补体,结合组织细胞等生物学活性。 功能区 H链和L链上每隔90个氨基酸残基通过链内二硫键连接形成跨度有110个氨基酸的环形结构域称为功能区,H链有VH,CH1,CH2,CH3等4个或5个功能区。IgG,IgA,IgD的H链有4个功能区,IgM和IgE的H链有5个功能区。L链有VL、CL两个功能区。 功能区的主要功能:
1)VH和VL抗原结合部位;2)CH1-3和CLIg遗传标志所在;3)CH2(IgG),CH3(IgM)C1q结合部位(补体结合位点);4)CH2—CH3(IgG)IgG可借该区主动通过胎盘由母体传给胎儿;5)CH3(IgG)FcγR结合部位;6)CH4(IgE)FcεR结合部位。 免疫球蛋白的水解片段木瓜蛋白酶水解片段
木瓜蛋白酶水解IgG的部位是在铰链区二硫键连接的2条重链的近N端,可将Ig裂解为两个完全相同的Fab段和一个Fc段。Fab段即抗原结合片段,相当于抗体分子的两个臂,由一条完整的轻链和重链的VH和CH1结构域组成。一个Fab片段为单价,可与抗原结合但不形成凝集反应或沉淀反应;Fc段即可结晶片段,相当于IgG的CH2和CH3结构域。Fc无抗原结合活性,是Ig与效应分子或细胞相互作用的部位。 免疫球蛋白的水解片段胃蛋白水解酶片段
胃蛋白酶作用于铰链区二硫键所连接的两条重链的近C端,水解Ig后可获得一个F(ab’)2片段和一些小片段pFc’。 F(ab’)2是由两个Fab及铰链区组成,由于Ig分子的两个臂仍由二硫键连接,因此F(ab’)2片段为双价,可同时结合两个抗原表位,故与抗原结合可发生凝集反应和沉淀反应,而且,由于F(ab’)2片段保留了结合相应抗原的生物学活性,又避免了Fc段免疫原性可能引起的副作用,因而被广泛用作生物制品。 免疫球蛋白的异质性类:在同一种属所有个体内,根据重链C区抗原特异性的差异,将重链分为5类,即免疫球蛋白分别为IgG,IgM,IgA,IgD和IgE。
亚类:同一类Ig,根据重链免疫反应性以及二硫键的数目和位置不问,又可分为若干个亚类。IgG有G1,G2,G3,G4四个亚类;IgA有A1,A2两个亚类;IgM有IgMl和IgM2两个亚类;IgD和IgE尚未发现亚类。
型:在同一种属所有个体内,根据Ig轻链C区抗原表位的差异,可将Ig轻链分为κ和λ,与此对应的免疫球蛋白分为κ和λ型。
亚型:同一型Ig中,根据轻链C区N端氨基酸排列的差异,又可将Ig分为亚型。例如λ链190氨基酸为亮氨酸时,称OZ(+);为精氨酸时,称OZ(-)。 外源因素所致的异质性—免疫球蛋白的多样性
多样性抗原的存在是导致免疫球蛋白异质性(即具有不同的抗原识别特异性)的外源因素,是免疫球蛋白异质性的物质基础。抗体的这种异质性,反映出机体对抗原精细结构的识别和应答。 内源因素所致的异质性—免疫球蛋白的血清型
免疫球蛋白既可以与相应的抗原发生特异性结合,其本身又可以激发机体产生特异性免疫应答,即免疫球蛋白也具有免疫原性,它之所以具有免疫原性是由于其分子中包含多种不同的抗原表位。免疫球蛋白分子上有三类不同的抗原表位,分别为同种型、同种异型和独特型抗原表位。那么采用血清学方法,可将Ig分为相应的血清型。 免疫球蛋白的功能IgG:是人和动物血清中含量少最高的免疫球蛋白,占血清免疫球蛋白总量的75%~80%。IgG是介导体液免疫的主要抗体,多以单体形式存在。它主要由脾脏和淋巴结中的浆细胞产生,大部分(45%~50%)存在于血浆中,其余存在于组织液和淋巴液中。IgG是惟一可以通过人(兔)胎盘的抗体,因此在新生儿的抗感染中起着十分重要的作用。IgG有四个亚类,IgG1、IgG2、IgG3、IgG4。IgG是动物自然感染和人工主动免疫后,机体所产生的主要抗体,它能调理、凝集和沉淀抗原,具有抗菌、抗病毒、抗毒素等免疫活性,在抗肿瘤免疫中,其作用也是不可缺少的;此外,它是引起II型、III型变态反应及自身免疫病的抗体,在肿瘤免疫中体内产生的封闭因子可能与IgG有关。IgM:是动物机体初次体液免疫反应最早产生的免疫球蛋白,其含量仅占血清免疫球蛋白的10%左右,主要由脾脏和淋巴结中B细胞产生,分布于血液中。IgM由5个单体组成的五聚体,又称为巨球蛋白。与IgG相比,IgM在体内产生最早,但持续时间短,因此不是机体抗感染免疫的主力,它在感染免疫的早期起着十分重要的作用,也可通过检测IgM抗体进行疫病的血清学早期诊断。IgM具有抗菌、抗病毒、中和毒素等免疫活性,它是一种高效能的抗体,其杀菌、溶菌、溶血、促进吞噬(调理作用)及凝集作用均比IgG高。IgM也有抗肿瘤作用,在补体参与下同样可介导对肿瘤细胞的破坏作用。此外,IgM可引起II型和III型变态反应及自身疫病而造成机体的损伤。IgA:以单体和二聚体两种分子形式存在,单体存在于血清中,称为血清型IgA,占血清免疫球蛋白的10%~20%;二聚体为分泌型IgA,是由呼吸道、消化道、泌尿生殖道等部位的黏膜固有层中的浆细胞所产生的。
分泌型IgA对机体呼吸道、消化道等局部黏膜免疫起着相当重要的作用,特别是对于一些经黏膜途径感染的病原微生物,黏膜免疫功能就显得十分重要,若动物机体呼吸道、消化道分泌液中存在这些病原微生物的相应的分泌型IgA抗体,则可抵抗其感染,因此分泌型IgA是机体黏膜免疫的一道“屏障”。IgE:是以单体分子形式存在,其重链比γ链多一个功能区(CH4),此区是与细胞结合的部位。IgE的产生部位与分泌型IgA相似,是上呼吸道和消化道黏膜固有层中的浆细胞所产生的,在血清中的含量甚微。IgE是一种亲细胞性抗体,其Fc片段中含有较多的半胱氨酸和蛋氨酸,这与其亲细胞性有关,因此IgE易于与皮肤组织、肥大细胞、血液中的嗜碱性粒细胞和血管内皮细胞结合。结合在肥大细胞和嗜碱性粒细胞上的IgE与抗原结合后,能引起这些细胞脱粒,释放组织胺等活性介质,从而引起I型过敏反应。IgE在抗寄生虫感染中具有重要的作用,诱导过敏反应,真菌感染后可诱导机体产生大量的IgE抗体。IgD:IgD很少分泌,在血清中的含量极低,而且极不稳定,容易降解。IgD主要作为成熟B细胞膜上的抗原特异性受体,是B细胞的重要表面标志,而且与免疫记忆有关。有报道认为IgD与某些过敏反应有关。 所有哺乳动物都具有4类主要的免疫球蛋白,即IgG、IgM、IgA和IgE。在所有种类动物中,每一类免疫球蛋白的基本特性没有明显的差异,但在其亚类和同种异型上存在一定的差异。 抗体产生的一般规律 抗体产生的一般规律包括初次应答、再次应答和回忆应答。
初次应答:某种抗原第一次进入动物机体引起的抗体产生的过程。主要抗体是IgM,然后是IgG;再次应答:在初次应答后几周或几月,当血液中抗体滴度很低,甚至消失后,同种抗原再次进入机体,引起的抗体产生过程。主要的抗体是IgG,其次是IgA;回忆应答: 抗原刺激机体产生的抗体经一定时间后,在体内逐渐消失,此时若机体再次接触相同的抗原物质,可使已消失的抗体快速回升的过程。主要抗体是IgG。 免疫球蛋白的生物学活性 特异性结合抗原
激活补体
结合FC段受体
通过胎盘和粘膜
免疫调节
2、什么是补体?
3、补体的经典途径如何?
4、补体激活后具有的生物学功能?第四章 特异性免疫
(6学时) 非典型肺炎流行猖獗引起社会的高度重视!但即使是流行最严重的地区,也不是人人都会得病!因为,冠状病毒虽然厉害,对于免疫功能强大的人,它也是无能为力的!目前全世界有大约6000万人感染了艾滋病毒,但从80年代中期以来,科学家发现,约有2%的病毒携带者竟然终身没有发展成艾滋病,成为“长期存活的病毒携带者”。这一现像表明,部份人对艾滋病毒具有先天的免疫力。津巴布韦一名6岁的艾滋病患儿第一道防线:屏障作用 人体的皮肤和黏膜等组成人体的第一道防线第二道防线:免疫吞噬细胞第三道防线:补体和溶菌酶等 特异性免疫应答是动物在个体发育过程中,由于受抗原异物的刺激,而发展生成有针对性的免疫力,包括引起生成致敏淋巴细胞的特异性细胞免疫和产生抗体所表现出的特异性体液免疫。这种免疫应答生成的原因,是由于机体与入侵的病原微生物或其他各种抗原异物相互斗争,或因接种疫苗,或因无意接触外界抗原异物后,引起T细胞及B细胞发生免疫应答而生成的。 特异性免疫的基本概念特异性免疫应答与非特异性免疫应答比较,它具有三个主要特点:①不能遗传给后代,它是在个体发育中,在非特异性免疫的基础上发展生成的,故属于后天获得性免疫。②具有严格的特异性,即它只对于引起其产生这种免疫力的同种抗原异物有对抗性,对于无关的抗原物质不起作用,故属于特异性免疫。③特异性免疫应答是在非特异性免疫应答的基础上建立起来的,也就是说,它需要非特异的体液因子及细胞因子相互配合协作,才能发挥更强大的免疫效能,既可表现免疫防护效果,又可引起免疫病理损害。 特异性免疫的基本过程 在抗原刺激下,机体形成特异性免疫应答,不论细胞免疫或体液免疫,都有其共同的规律,一般分为感应(识别)即加工处理抗原阶段、反应即淋巴细胞分化增殖阶段和效应即发生免疫应答(产生抗体或致敏淋巴细胞再次遇到抗原后刺激产生淋巴因子)三个阶段。1、感应阶段2、反应阶段3、效应阶段体液免疫过程抗原处理、呈递和识别。B细胞增殖分化,以及记忆细胞形成。抗体发挥免疫效应。体液免疫(抗原进入人体后被相应的抗体消灭)细胞免疫过程1、感应阶段2、反应阶段3、效应阶段抗原处理、呈递和识别。T细胞增殖分化,以及记忆细胞形成。效应T细胞、抗体和淋巴因子发挥免疫效应。细胞免疫(抗原进入人体后被相应的免疫细胞消灭) 淋巴细胞系(T细胞、B细胞、效应T细胞、效应B细胞)
吞噬细胞系(巨噬细胞,肥大细胞)。
粒细胞系(噬中性粒细胞、噬酸性粒细胞、噬碱性粒细胞)免疫细胞淋巴细胞的分化 实验探索强X光照射去 胸腺小鼠无免疫性输给小鼠来自胸腺的淋巴细胞 免疫功能得到恢复,但不能产生游离于体液中的抗体 输给来自骨髓的淋巴细胞 免疫功能不能恢复,但产生抗体的功能得到一定恢复 同时输入来自胸腺和骨髓的淋巴细胞小鼠就可恢复
全部免疫功能 实验结论: 胸腺内的淋巴细胞和骨髓中的淋巴细胞在免疫中起不同的作用,分别称为T细胞和B细胞。体液免疫细胞免疫体液免疫与细胞免疫的关系 两者各自有其独特作用,又可相互协作,共同发挥作用。各阶段注意点1、感应阶段: 吞噬细胞的作用是使抗原的抗原决定簇暴露,各细胞间的呈递是通过细胞的相互接触来完成的。2、反应阶段: 形成的记忆细胞在同种抗原再次侵入时,可迅速增殖、分化,形成大量的效应细胞,产生更强的特异性免疫反应。①体液免疫中,抗体与病菌结合后,可抑制其繁殖和对宿主细胞的黏附,防止感染和疾病的发生。抗体与病毒结合后,可使其失去侵染和破坏宿主细胞的能力。抗原和抗体结合后进一步的变化是形成沉淀或细胞集团,进而被吞噬细胞吞噬消化。3、效应阶段:②细胞免疫中,效应T细胞与被抗原入侵的宿主细胞(即靶细胞)接触,激活靶细胞的溶酶体酶,最终使靶细胞裂解死亡。细胞内的抗原也因失去藏身之所而为抗体消灭。 抗体抗体发现的过程:
19世纪末20世纪初,科学家们在实验中发现,用细菌或其外毒素给动物注射,过一段时间后,该动物的血清中出现一些有防御作用的保护性成分。
在20世纪30年代,科学家用电泳法鉴定,证明抗体是一种球蛋白。
1959年科学家对抗体的结构进行了研究,证明它是由四条肽链组成,借二硫键连接在一起,形成一个"Y"型的四链分子。不同的抗体某些部分氨基酸序列各不相同,决定抗体的特异性。 具有抗体活性的动物蛋白。主要存在于血浆中,也见于其他体液、组织和一些分泌液中。人血浆内的免疫球蛋白大多数存在于丙种球蛋白(γ-球蛋白)中。免疫球蛋白(Ig)Ig分子根据其可溶性、电荷、大小、化学结构等理化特性及其免疫反应性可分为IgA,IgG,IgD,IgE和IgM五类。在电镜下,IgG为“Y”型分子。“Y”的臂可结合抗原。若用化学方法处理免疫球蛋白,使其摆脱二硫键的束缚,分子就被裂解成4条独立的多肽链。所有Ig都具有由四条肽链组成的基本结构,称为单体(抗体单位),IgG为典型的单体。 抗体(antibody,Ab)是介导体液免疫的重要效应分子,是B细胞接受抗原刺激后增殖分化为浆细胞所产生的一种糖蛋白,主要存在于血清等体液中,能与相应抗原特异地结合,具有免疫功能。 抗体的概念 Porter对血清IgG抗体的研究证明,Ig分子基本结构是由四个肽链组成的,二条较小的轻链和二条较大的重链,轻链与重链是由二硫键连接形成,分为氨基端(N端),羧基端(C端)。任何一类天然免疫球蛋白分子均含有四条异源性多肽链,其中,相对分子质量较大的称为重链(heavy chain,H链),而相对分子质量较小的为轻链(light chain,L链)。同一天然Ig分子中的两条H链和两条L链的氨基酸组成完全相同。 重链和轻链重链:两条相同的长链称为重链、简称H链。H链由450—550氨基酸组成,相对分子质量为50—75kDa。根据Ig重链恒定区氨基酸的排列顺序不同,免疫反应性就不同,据此可将血清中的Ig分成五大类:IgG,IgA,IgM,IgD,IgE,其相应的重链分别为γ、μ、α、δ和ε链。 轻链:两条相同的短链称为轻链,简称L链,L链约由214个氨基酸组成,相对分子质量约为25kDa,根据L链免疫反应性不同,L链可分为两型,即κ型和λ型。根据λ链恒定区个别氨基酸的差异,又可分为λ1、λ2、λ3和λ4四个亚基(subtype)。同一天然的Ig分子上两条H链是同类,两条轻链是同型,五类Ig中,每一类Ig都可以有κ链或λ链,可以把Ig分成五类十型。 重链和轻链 可变区和恒定区 免疫球蛋白轻链和重链中靠近N端氨基酸序列变化较大的区域称为可变区(variable region,V区),分别占重链和轻链的1/4和1/2;而靠近C端氨基酸序列相对稳定的区域,称为恒定区(constant region,C区),分别占重链和轻链的3/4和1/2。可变区:免疫球蛋白轻链靠近氨基端(N端)的1/2区段或重链靠近氨基端(N端)1/5—1/4区段的110个氨基酸的组成和排列顺序在不同Ig中差异很大,称为可变区,分别用VH和VL表示。 可变区和恒定区 高变区(hypervariable region,HVR):在VH和VL中,各有3个区域的氨基酸组成和排列顺序高度可变,称为高变区或互补决定区(complementarity determining region,CDR),分别用HVRl(CDRl),HVR2(CDR2),HVR3(CDR3)表示。 恒定区:免疫球蛋白除了可变区以外,轻链靠近羧基端(C端)1/2区段或重链靠近C端3/4—4/5区段氨基酸的组成和排列相对稳定,称为恒定区。分别用CH和CL表示,不同Ig的CL的长度基本一致;不同Ig的CH长度不一,有的包括CH1,CH2,CH3或CH1,CH2,CH3,CH4。铰链区:位于CH1和CH2之间的非独立功能区,即位于Ig的Y形分子两臂和主干之间的区域,称为铰链区。富含脯氨酸,因此易伸展弯曲,易被木瓜蛋白酶、胃蛋白酶水解,并可通过变构有利于与不同距离的抗原决定簇结合。 可变区和恒定区铰链区的作用:
(1)当VL,VH与抗原结合时,此区发生扭曲,使抗体分子上两个抗原结合位点更好地与两个抗原决定簇发生互补,利于抗体分子高变区与抗原决定簇吻合。
(2)Ag和Ab结合后,铰链区发生构型变化,使存在于CH2区补体结合位点暴露,使补体得以结合,显示出活化补体,结合组织细胞等生物学活性。 功能区 H链和L链上每隔90个氨基酸残基通过链内二硫键连接形成跨度有110个氨基酸的环形结构域称为功能区,H链有VH,CH1,CH2,CH3等4个或5个功能区。IgG,IgA,IgD的H链有4个功能区,IgM和IgE的H链有5个功能区。L链有VL、CL两个功能区。 功能区的主要功能:
1)VH和VL抗原结合部位;2)CH1-3和CLIg遗传标志所在;3)CH2(IgG),CH3(IgM)C1q结合部位(补体结合位点);4)CH2—CH3(IgG)IgG可借该区主动通过胎盘由母体传给胎儿;5)CH3(IgG)FcγR结合部位;6)CH4(IgE)FcεR结合部位。 免疫球蛋白的水解片段木瓜蛋白酶水解片段
木瓜蛋白酶水解IgG的部位是在铰链区二硫键连接的2条重链的近N端,可将Ig裂解为两个完全相同的Fab段和一个Fc段。Fab段即抗原结合片段,相当于抗体分子的两个臂,由一条完整的轻链和重链的VH和CH1结构域组成。一个Fab片段为单价,可与抗原结合但不形成凝集反应或沉淀反应;Fc段即可结晶片段,相当于IgG的CH2和CH3结构域。Fc无抗原结合活性,是Ig与效应分子或细胞相互作用的部位。 免疫球蛋白的水解片段胃蛋白水解酶片段
胃蛋白酶作用于铰链区二硫键所连接的两条重链的近C端,水解Ig后可获得一个F(ab’)2片段和一些小片段pFc’。 F(ab’)2是由两个Fab及铰链区组成,由于Ig分子的两个臂仍由二硫键连接,因此F(ab’)2片段为双价,可同时结合两个抗原表位,故与抗原结合可发生凝集反应和沉淀反应,而且,由于F(ab’)2片段保留了结合相应抗原的生物学活性,又避免了Fc段免疫原性可能引起的副作用,因而被广泛用作生物制品。 免疫球蛋白的异质性类:在同一种属所有个体内,根据重链C区抗原特异性的差异,将重链分为5类,即免疫球蛋白分别为IgG,IgM,IgA,IgD和IgE。
亚类:同一类Ig,根据重链免疫反应性以及二硫键的数目和位置不问,又可分为若干个亚类。IgG有G1,G2,G3,G4四个亚类;IgA有A1,A2两个亚类;IgM有IgMl和IgM2两个亚类;IgD和IgE尚未发现亚类。
型:在同一种属所有个体内,根据Ig轻链C区抗原表位的差异,可将Ig轻链分为κ和λ,与此对应的免疫球蛋白分为κ和λ型。
亚型:同一型Ig中,根据轻链C区N端氨基酸排列的差异,又可将Ig分为亚型。例如λ链190氨基酸为亮氨酸时,称OZ(+);为精氨酸时,称OZ(-)。 外源因素所致的异质性—免疫球蛋白的多样性
多样性抗原的存在是导致免疫球蛋白异质性(即具有不同的抗原识别特异性)的外源因素,是免疫球蛋白异质性的物质基础。抗体的这种异质性,反映出机体对抗原精细结构的识别和应答。 内源因素所致的异质性—免疫球蛋白的血清型
免疫球蛋白既可以与相应的抗原发生特异性结合,其本身又可以激发机体产生特异性免疫应答,即免疫球蛋白也具有免疫原性,它之所以具有免疫原性是由于其分子中包含多种不同的抗原表位。免疫球蛋白分子上有三类不同的抗原表位,分别为同种型、同种异型和独特型抗原表位。那么采用血清学方法,可将Ig分为相应的血清型。 免疫球蛋白的功能IgG:是人和动物血清中含量少最高的免疫球蛋白,占血清免疫球蛋白总量的75%~80%。IgG是介导体液免疫的主要抗体,多以单体形式存在。它主要由脾脏和淋巴结中的浆细胞产生,大部分(45%~50%)存在于血浆中,其余存在于组织液和淋巴液中。IgG是惟一可以通过人(兔)胎盘的抗体,因此在新生儿的抗感染中起着十分重要的作用。IgG有四个亚类,IgG1、IgG2、IgG3、IgG4。IgG是动物自然感染和人工主动免疫后,机体所产生的主要抗体,它能调理、凝集和沉淀抗原,具有抗菌、抗病毒、抗毒素等免疫活性,在抗肿瘤免疫中,其作用也是不可缺少的;此外,它是引起II型、III型变态反应及自身免疫病的抗体,在肿瘤免疫中体内产生的封闭因子可能与IgG有关。IgM:是动物机体初次体液免疫反应最早产生的免疫球蛋白,其含量仅占血清免疫球蛋白的10%左右,主要由脾脏和淋巴结中B细胞产生,分布于血液中。IgM由5个单体组成的五聚体,又称为巨球蛋白。与IgG相比,IgM在体内产生最早,但持续时间短,因此不是机体抗感染免疫的主力,它在感染免疫的早期起着十分重要的作用,也可通过检测IgM抗体进行疫病的血清学早期诊断。IgM具有抗菌、抗病毒、中和毒素等免疫活性,它是一种高效能的抗体,其杀菌、溶菌、溶血、促进吞噬(调理作用)及凝集作用均比IgG高。IgM也有抗肿瘤作用,在补体参与下同样可介导对肿瘤细胞的破坏作用。此外,IgM可引起II型和III型变态反应及自身疫病而造成机体的损伤。IgA:以单体和二聚体两种分子形式存在,单体存在于血清中,称为血清型IgA,占血清免疫球蛋白的10%~20%;二聚体为分泌型IgA,是由呼吸道、消化道、泌尿生殖道等部位的黏膜固有层中的浆细胞所产生的。
分泌型IgA对机体呼吸道、消化道等局部黏膜免疫起着相当重要的作用,特别是对于一些经黏膜途径感染的病原微生物,黏膜免疫功能就显得十分重要,若动物机体呼吸道、消化道分泌液中存在这些病原微生物的相应的分泌型IgA抗体,则可抵抗其感染,因此分泌型IgA是机体黏膜免疫的一道“屏障”。IgE:是以单体分子形式存在,其重链比γ链多一个功能区(CH4),此区是与细胞结合的部位。IgE的产生部位与分泌型IgA相似,是上呼吸道和消化道黏膜固有层中的浆细胞所产生的,在血清中的含量甚微。IgE是一种亲细胞性抗体,其Fc片段中含有较多的半胱氨酸和蛋氨酸,这与其亲细胞性有关,因此IgE易于与皮肤组织、肥大细胞、血液中的嗜碱性粒细胞和血管内皮细胞结合。结合在肥大细胞和嗜碱性粒细胞上的IgE与抗原结合后,能引起这些细胞脱粒,释放组织胺等活性介质,从而引起I型过敏反应。IgE在抗寄生虫感染中具有重要的作用,诱导过敏反应,真菌感染后可诱导机体产生大量的IgE抗体。IgD:IgD很少分泌,在血清中的含量极低,而且极不稳定,容易降解。IgD主要作为成熟B细胞膜上的抗原特异性受体,是B细胞的重要表面标志,而且与免疫记忆有关。有报道认为IgD与某些过敏反应有关。 所有哺乳动物都具有4类主要的免疫球蛋白,即IgG、IgM、IgA和IgE。在所有种类动物中,每一类免疫球蛋白的基本特性没有明显的差异,但在其亚类和同种异型上存在一定的差异。 抗体产生的一般规律 抗体产生的一般规律包括初次应答、再次应答和回忆应答。
初次应答:某种抗原第一次进入动物机体引起的抗体产生的过程。主要抗体是IgM,然后是IgG;再次应答:在初次应答后几周或几月,当血液中抗体滴度很低,甚至消失后,同种抗原再次进入机体,引起的抗体产生过程。主要的抗体是IgG,其次是IgA;回忆应答: 抗原刺激机体产生的抗体经一定时间后,在体内逐渐消失,此时若机体再次接触相同的抗原物质,可使已消失的抗体快速回升的过程。主要抗体是IgG。 免疫球蛋白的生物学活性 特异性结合抗原
激活补体
结合FC段受体
通过胎盘和粘膜
免疫调节
同课章节目录
- 第一章 植物生命活动的调节
- 第一节 植物激素调节
- 第二节 其他调节
- 第二章 动物生命活动的调节
- 第一节 内环境与稳态
- 第二节 神经系统的结构与功能
- 第三节 高等动物的内分泌系统与体液调节
- 第三章 免疫系统与免疫功能
- 第一节 人体对抗病原体感染的非特异性防卫
- 第二节 特异性反应(免疫应答)
- 第三节 免疫系统的功能异常
- 第四章 种群
- 第一节 种群的特征
- 第二节 种群的增长方式
- 第三节 种群的数量波动及调节
- 第五章 群落
- 第一节 群落的物种组成和优势种
- 第二节 植物的生长型和群落结构
- 第三节 物种在群落中的生态位
- 第四节 群落的主要类型
- 第五节 群落演替
- 第六章 生态系统
- 第一节 生态系统的营养结构
- 第二节 生态系统中的生产量和生物量
- 第三节 能量流动和物质循环
- 第四节 生态系统的稳态及其调节
- 第七章 人类与环境
- 第一节 生物圈
- 第二节 全球人口动态
- 第三节 人类对全球环境的影响