浙科版必修2第二章第二节遗传的染色体学说(共95张PPT)
文档属性
| 名称 | 浙科版必修2第二章第二节遗传的染色体学说(共95张PPT) |

|
|
| 格式 | zip | ||
| 文件大小 | 4.4MB | ||
| 资源类型 | 教案 | ||
| 版本资源 | 浙科版 | ||
| 科目 | 生物学 | ||
| 更新时间 | 2014-10-18 00:00:00 | ||
图片预览







文档简介
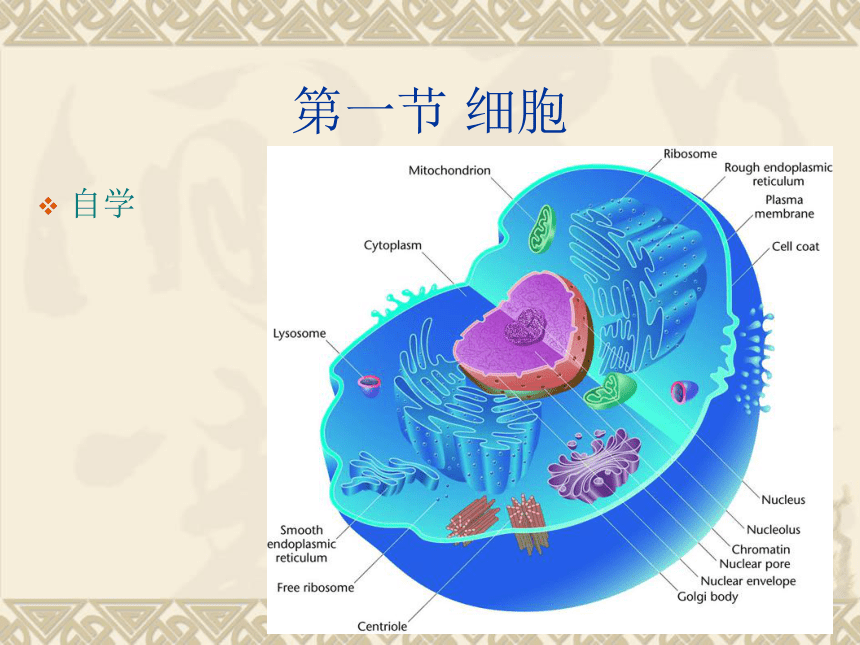
课件95张PPT。遗传的染色体学说 Chromosomal Theory of Heredity 第一节 细胞自学细胞:是由膜包围的,能进行独立繁殖的最小原生质团。
原生质:指细胞内所含的生活物质,真核细胞包括细胞膜、细胞质与细胞核。
细胞器:指存在于细胞中,利用光学或者电子显微镜可以分辨的、具有一定形态特点并执行特定功能的结构。一. 真核细胞的一般结构细胞的基本特征具有由磷脂双分子层与镶嵌蛋白构成的生物膜。
所有细胞都具有两种核酸:DNA与RNA。
所有细胞都具有蛋白质合成的机器-核糖体。
所有细胞都以一分为二的分裂方式进行增殖。
第二节 细胞分裂 2.1 染色质与染色体
2.2 染色体在有丝分裂中的行为
2.3 染色体在减数分裂中的行为
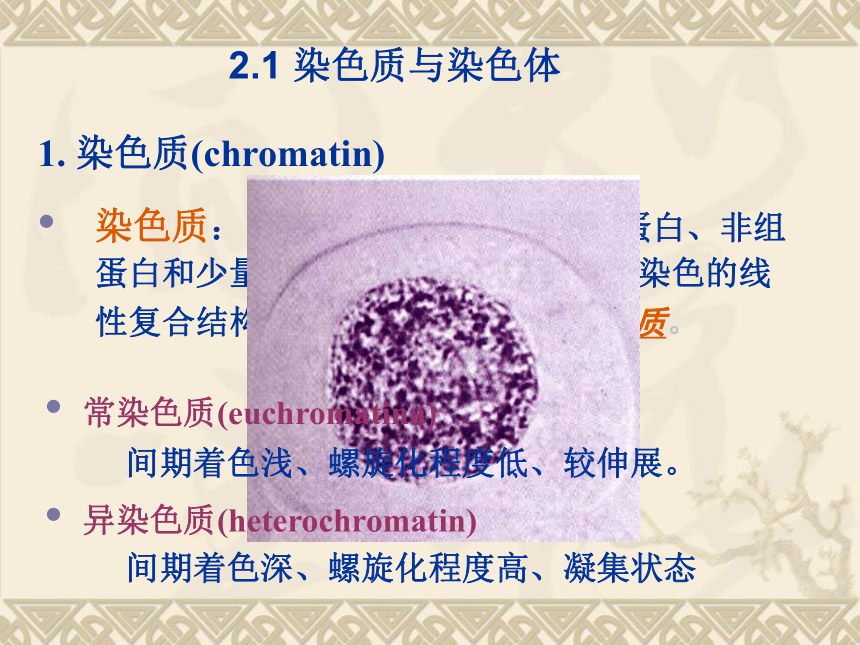
1. 染色质(chromatin)
染色质:间期细胞核内由DNA、组蛋白、非组蛋白和少量RNA组成、易被碱性染料染色的线性复合结构。包括常染色质和异染色质。 常染色质(euchromatina)
间期着色浅、螺旋化程度低、较伸展。
异染色质(heterochromatin)
间期着色深、螺旋化程度高、凝集状态 2.1 染色质与染色体 染色体(chromosome)则是染色质在细胞分裂过程中经过紧密缠绕、折叠、凝缩、精巧包装而成的具有固定形态的遗传物质存在形式,是高度螺旋化的DNA蛋白质纤维。 染色体的形态特征——随体
——次缢痕
——短臂
——主缢痕
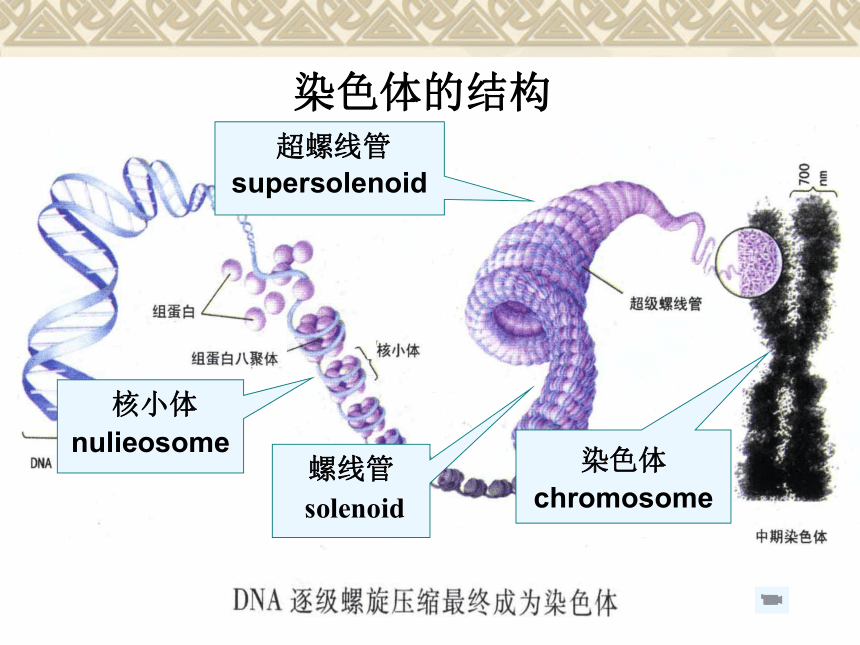
——长臂 染色质和染色体是真核生物遗传物质存在的两种不同形态,两者不存在成分上的差异,仅反映它们处于细胞分裂周期的不同功能阶段而已。 染色体的结构 核小体 nulieosome 超螺线管
supersolenoid
螺线管
solenoid染色体
chromosome 一级结构——核小体
二级结构——螺线体
三级结构——超螺线体
四级结构——中期染色体
?从DNA到染色体 DNA→核小体→螺线管→超螺线管→染色单体染色体的数目不同物种存在差异
同一物种相对恒定
体细胞中成双
生殖细胞中成单 水稻2n=24
玉米2n=20
陆地棉2n=52
萝卜2n=18
番茄2n=24染色体组型分析和显带 染色体组:一个生物赖以生存和保证正常生长与发育最低数目的一套染色体。
染色体组型:有丝分裂中期染色体的表现,包括一个生物种的染色体数目、形态和组成。
染色体显带:经过一系列处理和染色,使染色体出现染色深浅不同的区段。
(Q、G、R、C、N—带)
染色体组型分析(核型分析):对生物核内全部染色体的形态特征所进行的分析。正常男人的染色体组型及Giemsa显带光谱核型Spectral karyotyping,Sci,1996, 273:494
同源染色体(homologous chromosome)细胞中形态、结构和功能相同的一对染色体。(一个来自父方,一个来自母方)
非同源染色体(non-homologous chromosome) 细胞中形态、结构和功能不同的染色体互称为非同源染色体。
姊妹染色单体(sister chromatid):染色体通过复制形成的由同一着丝粒所连接在一起的两条遗传内容一样的子染色体。
非姊妹染色单体(non-sister chromatid) :同源染色体中来自父方染色体的单体与来自母方染色体的单体互称为非姊妹染色单体。特殊染色体多线染色体
双翅目昆虫(如果蝇)幼虫的唾腺细胞、肠细胞、 气管细胞等就发现多线染色体。
在幼虫发育期间, 染色体要进行4-15次复制,复制后的新老链不分离,因而一个染色体中就有24-215条DNA分子,果蝇幼虫唾腺细胞核中DNA的含量就比一般细胞多了1000多倍,这种只复制不分离的巨大染色体就称为多线染色体.由于多线染色体中平行排列的染色体纤维各段凝缩紧密程度不同,因而呈现出横行带纹现象.
果蝇唾腺多线染色体多线染色体的产生
多线染色体在有花植物中也有发现。有的植物,如百合,在合点细胞和反足细胞中也有巨大的多线染色体.植物多线染色体与动物多线染色体的不同 ①无明显的分带现象②染色单体间结合不紧密,各条单体清晰可见.
灯刷染色体
从上世纪末到本世纪初,自鱼类、爬行类、 鸟类以及某些无脊椎动物的卵母细胞中发现了另一类巨染色体,其特点是在中轴的两侧有许多精细而成对的环状突起,类似灯刷,因而称为灯刷染色体.
灯刷染色体(lampbrush chromosome)存在于动物卵母细胞中,是双线期4个染色单体;此期细胞合成mRNA和储备物质。爪蟾卵母细胞灯刷染色体含有上万个loops,虽然大部分DNA仍凝集在chromomeres,每个loop对应于一个特定的DNA序列。2. 2 染色体在有丝分裂中的行为 细胞的增殖是通过有丝分裂(mitosis)实现的,有丝分裂的结果是把一个细胞的整套染色体均等地分向两个子细胞,所以新形成的两个子细胞在遗传物质上跟原来的细胞是相同的。 2. 2. 1 细菌的有丝分裂 细菌属原核类(prokaryote)。细菌染色体位于细胞内的核区,核区外面没有核膜,所以称为原核。每一原核类细胞中通常只有一个染色体,染色体的结构简单,是一个裸露的DNA分子。2. 2. 1 细菌的有丝分裂 图 细菌细胞的有丝分裂 Color-enhanced electron micrograph of E.coli undergoing cell division. 2. 2. 2 真核类的有丝分裂因为真核类的细胞所含有的染色体数目往往较多,因而其细胞分裂也更为复杂。
有丝分裂过程是一个连续的过程,但为说明的方便起见,通常将其分成前期、中期、后期和末期四个时期,在两次有丝分裂中间的时期称为间期。 细胞周期(cell cycle)
上一次细胞分裂结束起到下一次细胞分裂止,细胞所经历的变化过程和时间。
例如:蚕豆根尖 19.5小时。
间期 分裂期
G1 — S — G2 — M
5 7.5 5 2
图 真核生物细胞周期示意图间期(interphase):光学显微镜下,细胞核是均匀一致的,看不到染色体,似乎静止 (活跃的代谢状态) 。
间期可分为三个时期:
G1期: DNA合成前期
S 期: DNA合成期
G2期: DNA合成后期(1) 前期(prophase): 间期核内的染色体细丝开始螺旋化,缩短变粗,染色体逐渐清晰起来。每一染色体含有纵向并列的两条染色单体,前期快结束时,染色体缩得很短。核膜核仁逐渐消失。
(2) 中期(metaphase): 中期开始时,核膜崩解,核质与胞质混和。纺锤体的细丝——纺锤丝与染色体的着丝粒区域连接。染色体向赤道面移动,着丝粒区域排列在赤道板上。这时最为容易计算染色体的数目。 纺锤丝出现,染色体聚集,
着丝点排列在赤道面上。(3) 后期(anaphase): 每一染色体的着丝粒已分裂为二,相互离开。着丝粒离开后,即被纺锤丝拉向两极,同时并列的染色单体也跟着分开,分别向两极移动。这时染色体又是单条了,也可叫做子染色体。 着丝点分开,染色单体分向两极(4) 末期(telophase)两组子染色体到达两极,染色体的螺旋结构逐渐消失,又出现核的重建过程。两个子核形成后,接着便发生细胞质的分割过程,把母细胞分隔成两个子细胞,细胞分裂结束(图)。 染色体到达两极且解螺旋,
子细胞、核仁核膜逐渐形成。有丝分裂的意义?有丝分裂过程中染色体的变迁:
从间期的S期?前期?中期,每个染色体具有两根染色单体(即具两条完整的DNA双链);从后期?末期?下一个细胞周期的G1期,在这些阶段中,所谓的染色体实质上只有一根染色单体(即只有一条DNA双链)。 2.3 染色体在减数分裂中的行为 减数分裂(meiosis)是一种特殊方式的细胞分裂,是在配子形成过程中发生的,包括两次连续的核分裂,但染色体只复制一次,因而在形成的四个子细胞核中,每个核只含有单倍数的染色体,即染色体数减少一半,所以把它叫做减数分裂。2. 3 染色体在减数分裂中的行为 两次连续的核分裂分别称为第一次减数分裂(减数分裂I,meiosis I)和第二次减数分裂(减数分裂II,meiosis II),在两次减数分裂中都能区分出前期、中期、后期和末期(图)。第一次分裂 前期I(prophaseI)
第一次减数分裂的前期特别长,包括细线期、偶线期、粗线期、双线期、终变期。
细线期
leptotene偶线期
zygotene粗线期
pachytene粗线期:染色体继续缩短变粗,两条同源染色体配对完毕。因此原来是2n条染色体,经配对后形成n组染色体,每一组合有2条同源染色体,这种配对的染色体叫做双价体(二价体)。双线期
diplotene双线期:双价体中的两条同源染色体开始分开,但分开不完全,并不形成两个独立的单价体,而是在两个同源染色体之间仍有若干处发生交叉而相互连接。交叉的地方实际上是染色单体发生了交换的结果终变期
diakinesis终变期:两条同源染色体仍有交叉联系着,所以仍为n个双价体。染色体变得更为粗短,螺旋化达到最高度,双价体开始向赤道面移动,分裂进入中期I。(2) 中期I: 各个双价体排列在赤道面上,两个同源染色体上的着丝粒逐渐远离,双价体开始分离,但仍有交叉联系着。 (3) 后期I: 双价体中的两条同源染色体分开,分别向两极移动,每一染色体有两个染色单体,在着丝粒区相连(相当于有丝分裂前期的一条染色体)。这样,每一极得到n条染色体,即在后期I时染色体数目减半。双价体中哪一条染色体移向哪一极是完全随机的。 (4) 末期I: 核膜重建,核仁重新形成,接着进行胞质分裂,成为两个子细胞。
注意:末期I的染色体只有n个,但每个染色体具有两条染色单体;而有丝分裂末期的染色体数为2n个,每个染色体只有一条染色单体。 (5) 减数间期: 在第一次分裂之末,两个子细胞进入间期,这时细胞核的形态与有丝分裂间期相似,但有许多生物没有间期,后期染色体直接进入第二次减数分裂的晚前期,染色体仍旧保持原来的浓缩状态。不过不论有没有间期,在两次减数分裂之间都没有DNA的合成及染色体的复制。 (6) 前期II、中期II、后期II和末期II 前期II、中期II、后期II和末期II的情况和有丝分裂过程完全一样,也是每一染色体具有两条染色单体,所不同的是染色体在第一次分裂过程中已经减数,只有n个染色体了。 第二次分裂前期II
prophase Ⅱ中期II
metophaseⅡ后期II
anaphaseⅡ末期II
telophaseⅡ四分体the tetrad二、 减数分裂的遗传学意义减数分裂是有性生殖生物产生性细胞所进行的细胞分裂方式;而两性性细胞受精结合(细胞融合)产生的合子是后代个体的起始点。
减数分裂不仅是生物有性繁殖必不可少的环节之一,也具有极为重要的遗传学意义。二、 减数分裂的遗传学意义保证了亲代与子代之间染色体数目的恒定性。
双亲性母细胞(2n)经过减数分裂产生性细胞(n),实现了染色体数目的减半;
雌雄性细胞融合产生的合子(及其所发育形成的后代个体)就具有该物种固有的染色体数目(2n),保持了物种的相对稳定。子代的性状遗传和发育得以正常进行。染色体数目的恒定二、 减数分裂的遗传学意义为生物的变异提供了重要的物质基础。
减数分裂中期 I,二价体的两个成员的排列方向是随机的,所以后期 I 分别来自双亲的两条同源染色体随机分向两极,因而所产生的性细胞就可能会有2n种非同源染色体的组合形式(染色体重组,recombination of chromosome)。如果某生物有两对同源染色体:AA’和BB’,产生的性细胞具有AA’中的一条和BB’中的一条。
非同源染色体在性细胞中可能有22=4种组合。中期 I 二价体的随机取向二、 减数分裂的遗传学意义为生物的变异提供了重要的物质基础。
另一方面,非姊妹染色单体间的交叉导致同源染色体间的片段交换(exchange of segment),使子细胞的遗传组成更加多样化,为生物变异提供更为重要的物质基础(染色体片断重组,recombination of segment)。同时这也是连锁遗传规律及基因连锁分析的基础。非姊妹染色单体交叉与片断交换
减数分裂与有丝分裂的异同点
名 称 间 期 前 期 中 期
有丝分裂 合成100%DNA 染色质→染色体 着丝点排列
赤道板上
减数分裂 间期Ⅰ,合成 前期Ⅰ,持续时间 中期Ⅰ,着
99.7%DNA 长,进行同源染色 丝点不排列
配对、联会、交 赤道板上,
换和交叉 。合 朝向两极
成0.3%DNA.
名 称 后 期 末 期 间 期Ⅱ
有丝分裂 着丝点分开 细胞质分裂形成 ————
染色单体分开 两个子细胞,染
色体数目不变
减数分裂 后期Ⅰ,着丝 末期Ⅰ,形成两个 短,无
点不分开,同 子细胞,染色体数 DNA合成
源染色体分开 目减半
第三节 染色体周史在所有以有性生殖繁殖的生物的生活史中,都会经历二倍体和单倍体的循环,只是它们在生活史中所占据的时间长短因生物的不同而异(图)。图 不同生物生活史中单倍体世代和二倍体世代所占的比例1. 动物的染色体周史 在动物中仅仅只是在减数分裂形成配子的阶段才存在单倍体,而在其他绝大部分的生活史中是以二倍体的形式存在的(图)。 图 动物的染色体周史 (以果蝇为例)精
子
形
成卵细胞形成2. 植物的染色体周史 植物的大部分生活史也是处在二倍体阶段,叫孢子体(sporophyte),只有一小部分是在单倍体阶段,叫配子体(gametophyte)。配子体只是经历极少数的几次有丝分裂(图)。 图 高等植物的染色体周史(以玉米为例)植物雌雄配子的形成植物的双受精:
卵细胞+精核 → 合子
2个极核+精核 → 3n胚乳 3. 真菌类的染色体周史 真菌的生活史是单倍体世代占优势,二倍体世代时间较短。
链孢霉(Neurospora crassa)有两种繁殖方式(图):一种是无性繁殖,当无性孢子(n)或菌丝落在营养物上,孢子萌发,菌丝生长形成菌丝体(n);另一种是有性繁殖,需要两个不同交配型(mating types)的菌丝体参与,一个交配型的单倍体核(n)通过另一相对交配型的子实体的受精丝,进入子实体中,核融合形成2n核(A/a)。脉孢霉只有在这个短暂时间内是二倍体世代。图 脉孢霉的染色体周史第四节 遗传的染色体学说1900年孟德尔定律的重新发现引起了人们对它的极大兴趣。1902年,美国W. S. Sutton和德国的T. Boveri各自独立地认识到豌豆产生配子时遗传因子的行为和性细胞在减数分裂过程中的染色体行为有着平行的关系:在真核生物中基因是成对存在的(等位基因),染色体也是成对存在的(同源染色体);
在形成配子时,等位基因相互分离,分别进入不同的配子中,一对同源染色体在第一次减数分裂时也相互分离,移向细胞的两极;
非等位基因在形成配子时,是自由组合地进入配子的,非同源染色体亦如此。因此,两位研究者各自独立地提出了细胞核的染色体可能是基因的载体的学说,即遗传的染色体学说(chromosome theory of heredity)。
按照这个学说,对孟德尔分离定律和自由组合定律可以做这样的理解:
在第一次减数分裂中,由于同源染色体的分离,使位于同源染色体上的等位基因分离,从而导致性状的分离(图);图 用染色体学说图解孟德尔分离定律当时,Sutton-Boveri这个假设引起了广泛的注意,因为这个假设不仅圆满地解释了孟德尔的两个定律,而且染色体是细胞中可见的结构,十分具体。虽然大多数人也认为这一假说极为合理,但并不是所有的生物学家都相信染色体上含有遗传信息,因为它缺乏真正的证据。要证实这个假设,进一步自然要把某一特定基因与特定染色体联系起来,将基因行为与在细胞分裂中染色体行为的平行关系转变为基因与染色体的从属关系。美国胚胎学家摩尔根在果蝇伴性遗传方面的发现及其基因理论的提出,以及他的学生Bridges的一系列工作使这一问题得到了很好的解决,这将在后面进—步学习。 有人观察到下列图像,请指出它们是有丝分裂还是减数分裂?母细胞染色体数是多少?减数分裂
前期Ⅱ
2n=6减数分裂
中期Ⅰ
2n=4有丝分裂
2n=4减数分裂
中期Ⅱ
2n=62 水稻的孢子体组织染色体数是12对,问下列各组织的染色体数目是多少?
胚乳 胚囊 叶片 种子胚
胚乳 3n=36
胚囊 3个反足细胞 2个助细胞, 2个极核, 1个卵细胞
8n=96
叶片 2n=24
种子胚 2n=24
3 人(2n=46)的(1)次级精母细胞,(2)精子细胞,(3)精子,(4)精原细胞(5)初级精母细胞(6)第一极体中各有多少条染色体?
(1)23
(2)23
(3)23
(4)46
(5)46
(6)234 某物种细胞的染色体数目为2n=24,分别说明下列各细胞分裂时期中有关数据:
(1)有丝分裂前期和后期染色体的着丝粒数;
(2)减数分裂前期I、后期I、前期II和后期II染色体着丝粒数;
(3)减数分裂前期I,中期I,末期I和后期II的染色体数。
(1)24, 48
(2)24,24,12,24
(3)24,24,12,24
原生质:指细胞内所含的生活物质,真核细胞包括细胞膜、细胞质与细胞核。
细胞器:指存在于细胞中,利用光学或者电子显微镜可以分辨的、具有一定形态特点并执行特定功能的结构。一. 真核细胞的一般结构细胞的基本特征具有由磷脂双分子层与镶嵌蛋白构成的生物膜。
所有细胞都具有两种核酸:DNA与RNA。
所有细胞都具有蛋白质合成的机器-核糖体。
所有细胞都以一分为二的分裂方式进行增殖。
第二节 细胞分裂 2.1 染色质与染色体
2.2 染色体在有丝分裂中的行为
2.3 染色体在减数分裂中的行为
1. 染色质(chromatin)
染色质:间期细胞核内由DNA、组蛋白、非组蛋白和少量RNA组成、易被碱性染料染色的线性复合结构。包括常染色质和异染色质。 常染色质(euchromatina)
间期着色浅、螺旋化程度低、较伸展。
异染色质(heterochromatin)
间期着色深、螺旋化程度高、凝集状态 2.1 染色质与染色体 染色体(chromosome)则是染色质在细胞分裂过程中经过紧密缠绕、折叠、凝缩、精巧包装而成的具有固定形态的遗传物质存在形式,是高度螺旋化的DNA蛋白质纤维。 染色体的形态特征——随体
——次缢痕
——短臂
——主缢痕
——长臂 染色质和染色体是真核生物遗传物质存在的两种不同形态,两者不存在成分上的差异,仅反映它们处于细胞分裂周期的不同功能阶段而已。 染色体的结构 核小体 nulieosome 超螺线管
supersolenoid
螺线管
solenoid染色体
chromosome 一级结构——核小体
二级结构——螺线体
三级结构——超螺线体
四级结构——中期染色体
?从DNA到染色体 DNA→核小体→螺线管→超螺线管→染色单体染色体的数目不同物种存在差异
同一物种相对恒定
体细胞中成双
生殖细胞中成单 水稻2n=24
玉米2n=20
陆地棉2n=52
萝卜2n=18
番茄2n=24染色体组型分析和显带 染色体组:一个生物赖以生存和保证正常生长与发育最低数目的一套染色体。
染色体组型:有丝分裂中期染色体的表现,包括一个生物种的染色体数目、形态和组成。
染色体显带:经过一系列处理和染色,使染色体出现染色深浅不同的区段。
(Q、G、R、C、N—带)
染色体组型分析(核型分析):对生物核内全部染色体的形态特征所进行的分析。正常男人的染色体组型及Giemsa显带光谱核型Spectral karyotyping,Sci,1996, 273:494
同源染色体(homologous chromosome)细胞中形态、结构和功能相同的一对染色体。(一个来自父方,一个来自母方)
非同源染色体(non-homologous chromosome) 细胞中形态、结构和功能不同的染色体互称为非同源染色体。
姊妹染色单体(sister chromatid):染色体通过复制形成的由同一着丝粒所连接在一起的两条遗传内容一样的子染色体。
非姊妹染色单体(non-sister chromatid) :同源染色体中来自父方染色体的单体与来自母方染色体的单体互称为非姊妹染色单体。特殊染色体多线染色体
双翅目昆虫(如果蝇)幼虫的唾腺细胞、肠细胞、 气管细胞等就发现多线染色体。
在幼虫发育期间, 染色体要进行4-15次复制,复制后的新老链不分离,因而一个染色体中就有24-215条DNA分子,果蝇幼虫唾腺细胞核中DNA的含量就比一般细胞多了1000多倍,这种只复制不分离的巨大染色体就称为多线染色体.由于多线染色体中平行排列的染色体纤维各段凝缩紧密程度不同,因而呈现出横行带纹现象.
果蝇唾腺多线染色体多线染色体的产生
多线染色体在有花植物中也有发现。有的植物,如百合,在合点细胞和反足细胞中也有巨大的多线染色体.植物多线染色体与动物多线染色体的不同 ①无明显的分带现象②染色单体间结合不紧密,各条单体清晰可见.
灯刷染色体
从上世纪末到本世纪初,自鱼类、爬行类、 鸟类以及某些无脊椎动物的卵母细胞中发现了另一类巨染色体,其特点是在中轴的两侧有许多精细而成对的环状突起,类似灯刷,因而称为灯刷染色体.
灯刷染色体(lampbrush chromosome)存在于动物卵母细胞中,是双线期4个染色单体;此期细胞合成mRNA和储备物质。爪蟾卵母细胞灯刷染色体含有上万个loops,虽然大部分DNA仍凝集在chromomeres,每个loop对应于一个特定的DNA序列。2. 2 染色体在有丝分裂中的行为 细胞的增殖是通过有丝分裂(mitosis)实现的,有丝分裂的结果是把一个细胞的整套染色体均等地分向两个子细胞,所以新形成的两个子细胞在遗传物质上跟原来的细胞是相同的。 2. 2. 1 细菌的有丝分裂 细菌属原核类(prokaryote)。细菌染色体位于细胞内的核区,核区外面没有核膜,所以称为原核。每一原核类细胞中通常只有一个染色体,染色体的结构简单,是一个裸露的DNA分子。2. 2. 1 细菌的有丝分裂 图 细菌细胞的有丝分裂 Color-enhanced electron micrograph of E.coli undergoing cell division. 2. 2. 2 真核类的有丝分裂因为真核类的细胞所含有的染色体数目往往较多,因而其细胞分裂也更为复杂。
有丝分裂过程是一个连续的过程,但为说明的方便起见,通常将其分成前期、中期、后期和末期四个时期,在两次有丝分裂中间的时期称为间期。 细胞周期(cell cycle)
上一次细胞分裂结束起到下一次细胞分裂止,细胞所经历的变化过程和时间。
例如:蚕豆根尖 19.5小时。
间期 分裂期
G1 — S — G2 — M
5 7.5 5 2
图 真核生物细胞周期示意图间期(interphase):光学显微镜下,细胞核是均匀一致的,看不到染色体,似乎静止 (活跃的代谢状态) 。
间期可分为三个时期:
G1期: DNA合成前期
S 期: DNA合成期
G2期: DNA合成后期(1) 前期(prophase): 间期核内的染色体细丝开始螺旋化,缩短变粗,染色体逐渐清晰起来。每一染色体含有纵向并列的两条染色单体,前期快结束时,染色体缩得很短。核膜核仁逐渐消失。
(2) 中期(metaphase): 中期开始时,核膜崩解,核质与胞质混和。纺锤体的细丝——纺锤丝与染色体的着丝粒区域连接。染色体向赤道面移动,着丝粒区域排列在赤道板上。这时最为容易计算染色体的数目。 纺锤丝出现,染色体聚集,
着丝点排列在赤道面上。(3) 后期(anaphase): 每一染色体的着丝粒已分裂为二,相互离开。着丝粒离开后,即被纺锤丝拉向两极,同时并列的染色单体也跟着分开,分别向两极移动。这时染色体又是单条了,也可叫做子染色体。 着丝点分开,染色单体分向两极(4) 末期(telophase)两组子染色体到达两极,染色体的螺旋结构逐渐消失,又出现核的重建过程。两个子核形成后,接着便发生细胞质的分割过程,把母细胞分隔成两个子细胞,细胞分裂结束(图)。 染色体到达两极且解螺旋,
子细胞、核仁核膜逐渐形成。有丝分裂的意义?有丝分裂过程中染色体的变迁:
从间期的S期?前期?中期,每个染色体具有两根染色单体(即具两条完整的DNA双链);从后期?末期?下一个细胞周期的G1期,在这些阶段中,所谓的染色体实质上只有一根染色单体(即只有一条DNA双链)。 2.3 染色体在减数分裂中的行为 减数分裂(meiosis)是一种特殊方式的细胞分裂,是在配子形成过程中发生的,包括两次连续的核分裂,但染色体只复制一次,因而在形成的四个子细胞核中,每个核只含有单倍数的染色体,即染色体数减少一半,所以把它叫做减数分裂。2. 3 染色体在减数分裂中的行为 两次连续的核分裂分别称为第一次减数分裂(减数分裂I,meiosis I)和第二次减数分裂(减数分裂II,meiosis II),在两次减数分裂中都能区分出前期、中期、后期和末期(图)。第一次分裂 前期I(prophaseI)
第一次减数分裂的前期特别长,包括细线期、偶线期、粗线期、双线期、终变期。
细线期
leptotene偶线期
zygotene粗线期
pachytene粗线期:染色体继续缩短变粗,两条同源染色体配对完毕。因此原来是2n条染色体,经配对后形成n组染色体,每一组合有2条同源染色体,这种配对的染色体叫做双价体(二价体)。双线期
diplotene双线期:双价体中的两条同源染色体开始分开,但分开不完全,并不形成两个独立的单价体,而是在两个同源染色体之间仍有若干处发生交叉而相互连接。交叉的地方实际上是染色单体发生了交换的结果终变期
diakinesis终变期:两条同源染色体仍有交叉联系着,所以仍为n个双价体。染色体变得更为粗短,螺旋化达到最高度,双价体开始向赤道面移动,分裂进入中期I。(2) 中期I: 各个双价体排列在赤道面上,两个同源染色体上的着丝粒逐渐远离,双价体开始分离,但仍有交叉联系着。 (3) 后期I: 双价体中的两条同源染色体分开,分别向两极移动,每一染色体有两个染色单体,在着丝粒区相连(相当于有丝分裂前期的一条染色体)。这样,每一极得到n条染色体,即在后期I时染色体数目减半。双价体中哪一条染色体移向哪一极是完全随机的。 (4) 末期I: 核膜重建,核仁重新形成,接着进行胞质分裂,成为两个子细胞。
注意:末期I的染色体只有n个,但每个染色体具有两条染色单体;而有丝分裂末期的染色体数为2n个,每个染色体只有一条染色单体。 (5) 减数间期: 在第一次分裂之末,两个子细胞进入间期,这时细胞核的形态与有丝分裂间期相似,但有许多生物没有间期,后期染色体直接进入第二次减数分裂的晚前期,染色体仍旧保持原来的浓缩状态。不过不论有没有间期,在两次减数分裂之间都没有DNA的合成及染色体的复制。 (6) 前期II、中期II、后期II和末期II 前期II、中期II、后期II和末期II的情况和有丝分裂过程完全一样,也是每一染色体具有两条染色单体,所不同的是染色体在第一次分裂过程中已经减数,只有n个染色体了。 第二次分裂前期II
prophase Ⅱ中期II
metophaseⅡ后期II
anaphaseⅡ末期II
telophaseⅡ四分体the tetrad二、 减数分裂的遗传学意义减数分裂是有性生殖生物产生性细胞所进行的细胞分裂方式;而两性性细胞受精结合(细胞融合)产生的合子是后代个体的起始点。
减数分裂不仅是生物有性繁殖必不可少的环节之一,也具有极为重要的遗传学意义。二、 减数分裂的遗传学意义保证了亲代与子代之间染色体数目的恒定性。
双亲性母细胞(2n)经过减数分裂产生性细胞(n),实现了染色体数目的减半;
雌雄性细胞融合产生的合子(及其所发育形成的后代个体)就具有该物种固有的染色体数目(2n),保持了物种的相对稳定。子代的性状遗传和发育得以正常进行。染色体数目的恒定二、 减数分裂的遗传学意义为生物的变异提供了重要的物质基础。
减数分裂中期 I,二价体的两个成员的排列方向是随机的,所以后期 I 分别来自双亲的两条同源染色体随机分向两极,因而所产生的性细胞就可能会有2n种非同源染色体的组合形式(染色体重组,recombination of chromosome)。如果某生物有两对同源染色体:AA’和BB’,产生的性细胞具有AA’中的一条和BB’中的一条。
非同源染色体在性细胞中可能有22=4种组合。中期 I 二价体的随机取向二、 减数分裂的遗传学意义为生物的变异提供了重要的物质基础。
另一方面,非姊妹染色单体间的交叉导致同源染色体间的片段交换(exchange of segment),使子细胞的遗传组成更加多样化,为生物变异提供更为重要的物质基础(染色体片断重组,recombination of segment)。同时这也是连锁遗传规律及基因连锁分析的基础。非姊妹染色单体交叉与片断交换
减数分裂与有丝分裂的异同点
名 称 间 期 前 期 中 期
有丝分裂 合成100%DNA 染色质→染色体 着丝点排列
赤道板上
减数分裂 间期Ⅰ,合成 前期Ⅰ,持续时间 中期Ⅰ,着
99.7%DNA 长,进行同源染色 丝点不排列
配对、联会、交 赤道板上,
换和交叉 。合 朝向两极
成0.3%DNA.
名 称 后 期 末 期 间 期Ⅱ
有丝分裂 着丝点分开 细胞质分裂形成 ————
染色单体分开 两个子细胞,染
色体数目不变
减数分裂 后期Ⅰ,着丝 末期Ⅰ,形成两个 短,无
点不分开,同 子细胞,染色体数 DNA合成
源染色体分开 目减半
第三节 染色体周史在所有以有性生殖繁殖的生物的生活史中,都会经历二倍体和单倍体的循环,只是它们在生活史中所占据的时间长短因生物的不同而异(图)。图 不同生物生活史中单倍体世代和二倍体世代所占的比例1. 动物的染色体周史 在动物中仅仅只是在减数分裂形成配子的阶段才存在单倍体,而在其他绝大部分的生活史中是以二倍体的形式存在的(图)。 图 动物的染色体周史 (以果蝇为例)精
子
形
成卵细胞形成2. 植物的染色体周史 植物的大部分生活史也是处在二倍体阶段,叫孢子体(sporophyte),只有一小部分是在单倍体阶段,叫配子体(gametophyte)。配子体只是经历极少数的几次有丝分裂(图)。 图 高等植物的染色体周史(以玉米为例)植物雌雄配子的形成植物的双受精:
卵细胞+精核 → 合子
2个极核+精核 → 3n胚乳 3. 真菌类的染色体周史 真菌的生活史是单倍体世代占优势,二倍体世代时间较短。
链孢霉(Neurospora crassa)有两种繁殖方式(图):一种是无性繁殖,当无性孢子(n)或菌丝落在营养物上,孢子萌发,菌丝生长形成菌丝体(n);另一种是有性繁殖,需要两个不同交配型(mating types)的菌丝体参与,一个交配型的单倍体核(n)通过另一相对交配型的子实体的受精丝,进入子实体中,核融合形成2n核(A/a)。脉孢霉只有在这个短暂时间内是二倍体世代。图 脉孢霉的染色体周史第四节 遗传的染色体学说1900年孟德尔定律的重新发现引起了人们对它的极大兴趣。1902年,美国W. S. Sutton和德国的T. Boveri各自独立地认识到豌豆产生配子时遗传因子的行为和性细胞在减数分裂过程中的染色体行为有着平行的关系:在真核生物中基因是成对存在的(等位基因),染色体也是成对存在的(同源染色体);
在形成配子时,等位基因相互分离,分别进入不同的配子中,一对同源染色体在第一次减数分裂时也相互分离,移向细胞的两极;
非等位基因在形成配子时,是自由组合地进入配子的,非同源染色体亦如此。因此,两位研究者各自独立地提出了细胞核的染色体可能是基因的载体的学说,即遗传的染色体学说(chromosome theory of heredity)。
按照这个学说,对孟德尔分离定律和自由组合定律可以做这样的理解:
在第一次减数分裂中,由于同源染色体的分离,使位于同源染色体上的等位基因分离,从而导致性状的分离(图);图 用染色体学说图解孟德尔分离定律当时,Sutton-Boveri这个假设引起了广泛的注意,因为这个假设不仅圆满地解释了孟德尔的两个定律,而且染色体是细胞中可见的结构,十分具体。虽然大多数人也认为这一假说极为合理,但并不是所有的生物学家都相信染色体上含有遗传信息,因为它缺乏真正的证据。要证实这个假设,进一步自然要把某一特定基因与特定染色体联系起来,将基因行为与在细胞分裂中染色体行为的平行关系转变为基因与染色体的从属关系。美国胚胎学家摩尔根在果蝇伴性遗传方面的发现及其基因理论的提出,以及他的学生Bridges的一系列工作使这一问题得到了很好的解决,这将在后面进—步学习。 有人观察到下列图像,请指出它们是有丝分裂还是减数分裂?母细胞染色体数是多少?减数分裂
前期Ⅱ
2n=6减数分裂
中期Ⅰ
2n=4有丝分裂
2n=4减数分裂
中期Ⅱ
2n=62 水稻的孢子体组织染色体数是12对,问下列各组织的染色体数目是多少?
胚乳 胚囊 叶片 种子胚
胚乳 3n=36
胚囊 3个反足细胞 2个助细胞, 2个极核, 1个卵细胞
8n=96
叶片 2n=24
种子胚 2n=24
3 人(2n=46)的(1)次级精母细胞,(2)精子细胞,(3)精子,(4)精原细胞(5)初级精母细胞(6)第一极体中各有多少条染色体?
(1)23
(2)23
(3)23
(4)46
(5)46
(6)234 某物种细胞的染色体数目为2n=24,分别说明下列各细胞分裂时期中有关数据:
(1)有丝分裂前期和后期染色体的着丝粒数;
(2)减数分裂前期I、后期I、前期II和后期II染色体着丝粒数;
(3)减数分裂前期I,中期I,末期I和后期II的染色体数。
(1)24, 48
(2)24,24,12,24
(3)24,24,12,24
同课章节目录
- 前言
- 第一章 孟德尔定律
- 第一节 分离定律
- 第二节 自由组合定律
- 第二章 染色体与遗传
- 第一节 减数分裂中的染色体行为
- 第二节 遗传的染色体学说
- 第三节 性染色体与伴性遗传
- 第三章 遗传的分子基础
- 第一节 核酸是遗传物质的证据
- 第二节 DNA的分子结构和特点
- 第三节 遗传信息的传递
- 第四节 遗传信息的表达—-RNA和蛋白质的合成
- 第四章 生物的变异
- 第一节 生物变异的来源
- 第二节 生物变异在生产上的应用
- 第五章 生物的进化
- 第一节 生物的多样性、统一性和进化
- 第二节 进化性变化是怎样发生的
- 第三节 探索生物进化的历史
- 第六章 遗传与人类健康
- 第一节 人类遗传病的主要类型
- 第二节 遗传咨询与优生
- 第三节 基因治疗和人类基因组计划
- 第四节 遗传病与人类未来