人教版必修一第六章第三节细胞衰老与凋亡(共65张PPT)
文档属性
| 名称 | 人教版必修一第六章第三节细胞衰老与凋亡(共65张PPT) |

|
|
| 格式 | zip | ||
| 文件大小 | 15.3MB | ||
| 资源类型 | 教案 | ||
| 版本资源 | 人教版(新课程标准) | ||
| 科目 | 生物学 | ||
| 更新时间 | 2014-11-17 00:00:00 | ||
图片预览










文档简介
课件65张PPT。CELL BIOLOGY细 胞 生 物 学C第一章 绪论
第二章 细胞基本知识概要
第三章 细胞生物学研究方法
第四章 细胞质膜与细胞表面
第五章 物质跨膜运输与信号传递
第六章 细胞质基质与内膜系统
第七章 细胞的能量转换
第八章 细胞核与染色体
第九章 核糖体
第 十 章 细胞骨架
第十一章 细胞增殖及其调控
第十二章 细胞分化与基因表达调控
第十三章 细胞衰老与凋亡
第十三章 细胞衰老和凋亡第一节 细胞衰老(cellular aging或cell senescence)又称老化,通常指生物发育成熟后,在正常情况下随着年龄的增加,机能减退,内环境稳定性下降,结构中心组分退行性变化,趋向死亡的不可逆的现象。
衰老和死亡是生命的基本现象,衰老过程发生在生物界的整体水平、种群水平、个体水平、细胞水平以及分子水平等不同的层次。一、Hayflick 界限(Hayflick Limitation)● Hayflick 界限:细胞,至少是培养的细胞,不是不死的,而是有一定的寿命;它们的增殖能力不是无限的,而是有一定的界限。“不死性”学说的错误在于:
◆癌细胞或培养的细胞系是不正常细胞,其染色体数目或形态已经不同于原先的细胞。
◆细胞的增殖能力与供体年龄有关。
◆物种寿命与培养细胞寿命之间存在着一定的关系。
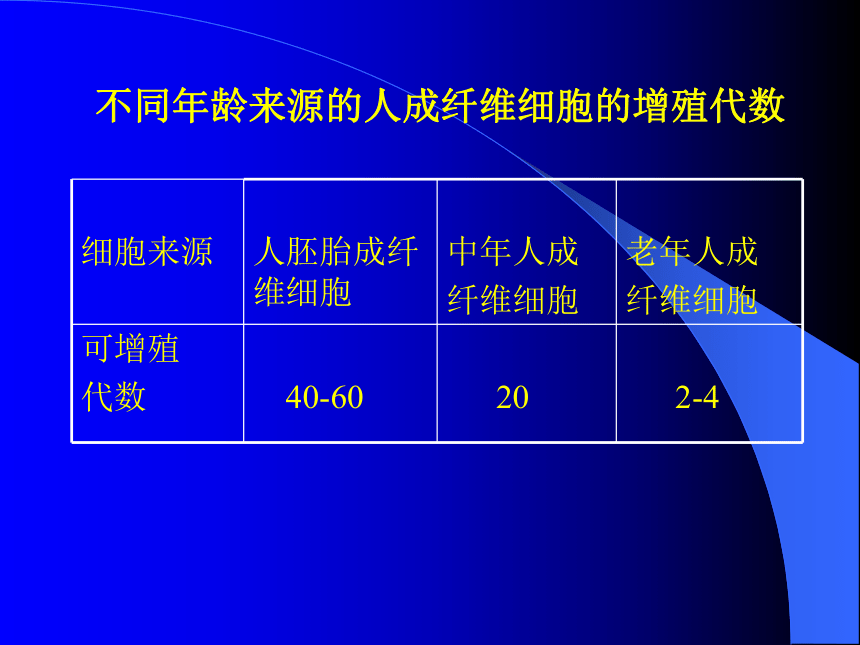
◆衰老的原因在于细胞本身。不同年龄来源的人成纤维细胞的增殖代数二、细胞在体内条件下的衰老●在机体内,细胞的衰老和死亡是常见的现象,甚至在个体发育的早期也会发生。
●正常情况下终生保持分裂的细胞,其分裂能力是否随着有机体年龄的增高而下降?它们 会不会衰老?
◆衰老动物体内,细胞分裂速度显著减慢,其原因主要是G1期明
显延长。
◆衰老个体内的环境因素影响了细胞的增殖和衰老。
◆骨髓干细胞移植实验说明随着年龄的增加,干细胞增殖速度也
趋缓慢。三、衰老细胞结构的变化●细胞核的变化:核膜内折,染色质固缩化。
●内质网的变化: 衰老动物内质网成分弥散性地分散于核周胞质中,粗面内质网的总量似乎是减少了。
●线粒体的变化:通常,细胞中线粒体的数量随年龄增大而减少,而其体积则随年龄增大而增大。
●致密体的生成:由溶酶体和线粒体转化而来。
●膜系统的变化
◆衰老的细胞,其膜流动性降低、韧性减小。
◆衰老细胞间间隙连接及膜内颗粒的分布也发生变化。(一)形态变化核 增大、染色深、核内有包含物
染色质 凝聚、固缩、碎裂、溶解
质膜 粘度增加、流动性降低
细胞质 色素积聚、空泡形成
线粒体 数目减少、体积增大、mt DNA突变或丢失
高尔基体 碎裂
尼氏体 消失
包含物 糖原减少、脂肪积聚
核膜 内陷表15-1 衰老细胞的形态变化尼氏体:围绕某些感觉神经末梢的卵圆形同心层状结构 。1、DNA:复制与转录受阻,端粒DNA、mtDNA缺失。DNA氧化、断裂、缺失和交联,甲基化程度降低。
2、RNA:m RNA和t RNA含量降低。
3、蛋白质:含成下降,细胞内蛋白质发生糖基化、氨甲酰化、脱氨基等修饰反应,导致蛋白质稳定性、抗原性,可消化性下降,自由基使蛋白质肽断裂,交联而变性。氨基酸由左旋变为右旋。
4、酶分子:活性中心被氧化,金属离子丢失,酶分子的二级结构,溶解度,等电点发生改变,酶失活。
5、脂类:不饱和脂肪酸被氧化。引起膜脂之间或与脂蛋白之间交联,膜的流动性降低。 (二)分子水平的变化四、细胞衰老的分子机理 该理论认为,代谢过程中产生的活性氧基团或分子(reactive oxygen species,ROS)引发的氧化性损伤的积累,最终导致衰老。
自由基:是一类瞬时形成的含不成对电子的原子或功能基团,普遍存在于生物系统。具有高度反应活性,可引发链式自由基反应,引起DNA、蛋白质和脂类,尤其是多不饱和脂肪酸等大分子物质变性和交联,损伤DNA、生物膜、重要的结构蛋白和功能蛋白,从而引起衰老各种现象的发生。
ROS主要有三种类型:
① O2-,即超氧自由基; ② OH-,即羟自由基; ③ H2O2。
(一)氧化性损伤学说:内源性自由基是人体自由基的主要来源,其产生的主要途径有:
①由线粒体呼吸链电子泄漏产生;
②由经过氧化物酶体的多功能氧化酶(MFO)等催化底物羟化产生。
③机体血红蛋白、肌红蛋白中还可通过非酶促反应产生自由基。
清除ROS,就可以延长寿命。正常细胞内存在清除自由基的防御系统,包括酶系统和非酶系统:
酶系统:SOD,CAT,GR等;
非酶系统:维生素E,醌类等电子受体。
(二)端粒与衰老: 细胞增殖次数与端粒DNA长度有关。
体细胞染色体的端粒DNA会随细胞分裂次数增加而不断缩短。细胞DNA每复制一次端粒就缩短一段,当缩短到一定程度至Hayflick点时,可能会启动DNA损伤检测点(DNA damage checkpoint),激活p53,引起p21表达,导致不可逆地退出细胞周期,走向衰亡。资料表明人的成纤维细胞端粒每年缩短14-18bp,可见染色体的端粒有细胞分裂计数器的功能,能记忆细胞分裂的次数。
端粒的长度与端粒酶(telomerase)的活性有关。
端粒酶是一种反转录酶,能以自身的RNA为模板合成端粒DNA,在精原细胞、干细胞和肿瘤细胞(如Hela细胞)中有较高的端粒酶活性,而正常体细胞中端粒酶的活性很低,呈抑制状态。(三)r DNA与衰老
在S.cerevisiae该酵母细胞中,会通过同源重组形成染色体外rDNA环
(extrachromosomal rDNA circle, ESC)。当ESC积累到500~1000拷贝是,细胞就进入衰老。但在哺乳动物中ESC并不多见。
(四)沉默信息调节蛋白(Sir )复合物与衰老
Sir复合物的功能是阻值它们所在位点的 DNA 转录。
(五)SGS1基因、WRN基因与衰老
SGS1基因编码的蛋白属于RecQ亚族的DNA解旋酶。WRN基因与SGS1基因同源,属于DNA解旋酶。在Werner综合症(WS)的早衰患者中观察不到WRN蛋白。Werner's syndrome 成人早衰症(Werner's syndrome)病人平均39岁时出现衰老,47岁左右生命结束,患婴幼儿早衰症(Hutchinson-Gilford syndrome,图15-2)的小孩在1岁时出现明显的衰老,12-18岁即过早夭折。
(六)程序性衰老(programmed senescence) 程序性衰老理论认为:生物的生长、发育、衰老和死亡都由基因程序控制的,衰老实际上是某些基因依次开启或关闭的结果。
例如在小鼠肝中,胚胎早期表达的胞质丙氨酸转氨酶(cytosolic alanine aminotransferase,c AAT)为 A 型,随后停止表达,但是在衰老时则表达 B 型 c AAT,其它类似的衰老标志物(senescence markers)也有报道,如肝脏中的衰老标志蛋白 2(senescence marker protein 2)也是在老年期表达。
(七)线粒体 DNA 突变(mitochondrial DNA mutation) 在线粒体氧化磷酸化生成 ATP 的过程中,大约有1-4%氧转化ROS,因此线粒体是自由基浓度最高的细胞器。 mtDNA裸露于基质,缺乏结合蛋白的保护,最易受自由基伤害,而催化mtDNA复制的DNA聚合酶γ不具有校正功能,复制错误频率高,同时缺乏有效的修复酶,故mtDNA最容易发生突变。Mt DNA突变使呼吸链功能受损,进一步引起自由基堆积,如此反复循环。衰老个体细胞中mtDNA缺失表现明显,并随着年龄的增加而增加,许多研究认为 mtDNA缺失与衰老及伴随的老年衰退性疾病有密切关系。
人类的脑、心、骨骼肌的氧负荷(oxidative stress)最大,因而是最容易衰老的组织。研究表明,限制热量(caloric restriction)摄入能明显延长小鼠的寿命。第二节 细胞凋亡 (Apoptosis)一、细胞凋亡的概念及其生物学意义● 概念:
细胞凋亡是一个主动的由基因决定的自动结束生命的过程,也常常被称为细胞程序死亡(programmed cell death, PCD)。凋亡细胞将被吞噬细胞吞噬。
● 细胞调亡的生物学意义:
它是维持组织机能和形态所必需的。生物发育过程中及成体组织中正常的细胞凋亡有助于保证细胞只在需要它们的时候和需要它们活的地方存活。这对于多细胞生物个体发育的正常进行,自稳平衡的保持以及抵御外界各种因素 的干扰方面都起着非常关键的作用。 蝌蚪尾的消失
发育过程中手和足
的成形过程 脊椎动物的神经系统的发育 脊椎动物的神经
系统的发育 二、细胞凋亡的形态学和生物化学特征 细胞死亡的方式通常有3种:即①细胞坏死(necrosis)。②细胞凋亡(apoptosis)。③细胞程序性死亡(programmed cell death,PCD)。
(一)细胞坏死
是细胞受到化学因素(如强酸、强碱、有毒物质)、物理因素(如热、辐射)和生物因素(如病原体)等环境因素的伤害,引起细胞死亡的现象。
早期表现为胞质内线粒体和内质网肿胀、崩解,核发生固缩或断裂。随着胞质内蛋白变性、凝固或碎裂,以及嗜碱性核蛋白的降解,细胞质呈现强嗜酸性,故坏死组织或细胞在苏木精/伊红染色切片中,胞质呈均一的深伊红色,原有的微细结构消失。
在含水量高的细胞,可因胞质内水泡不断增大,并发生溶解,导致细胞结构完全消失,最后细胞膜和细胞器破裂,DNA降解,细胞内容物流出,引起周围组织炎症反应。(二)细胞凋亡的形态学特征Kerr( 1972)最先提出,与细胞坏死的区别是:
①染色质聚集、分块、位于核膜上,胞质凝缩,最后核断裂,细胞通过出芽的方式形成许多凋亡小体(apoptotic bodies);
②凋亡小体内有结构完整的细胞器,还有凝缩的染色体,可被邻近细胞吞噬消化,因始终有膜封闭,没有内溶物释放,故不会引起炎症;
③凋亡细胞中仍需要合成一些蛋白质,但是在坏死细胞中ATP和蛋白质合成受阻或终止;
④核酸内切酶活化,导致染色质 DNA 在核小体连接部位断裂,形成约 200bp 整数倍的核酸片段,凝胶电泳图谱呈梯状;
⑤凋亡通常是生理性变化,而细胞坏死是病理性变化。
表15-2 细胞凋亡和细胞坏死的区别区别点 细胞凋亡 细胞坏死
起因 生理或病理性 病理性变化或剧烈损伤
范围 单个散在细胞 大片组织或成群细胞
细胞膜 保持完整,一直到形成凋亡小体 破损
染色质 凝聚在核膜下呈半月状 呈絮状
细胞器 无明显变化 肿胀、内质网崩解
细胞体积 固缩变小 肿胀变大
凋亡小体 有,被邻近细胞或 无,细胞自溶,残余碎片
巨噬细胞吞噬 被巨噬细胞吞噬
基因组DNA 有控降解,电泳图谱呈梯状 随机降解,电泳图谱呈涂抹状
蛋白质合成 有 无
调节过程 受基因调控 被动进行
炎症反应 无,不释放细胞内容物 有,释放内容物。细胞坏死与凋亡的形态区别坏死细胞凋亡细胞细胞凋亡的形态学特征? Morphologi changes:
Early : Chromosome condensation, cell body shrink
Later : Blebbing and Nucleus and cytoplasm fragment—Apoptotic bodies
At last: Phagocytosed弯曲胸腺细胞正常凋亡正常凋亡(三)细胞凋亡的生化特征核酸内切酶活化,导致染色质DNA在核小体连接部位断裂,形成约200bp整数倍的核酸片段,凝胶电泳图谱呈梯状
细胞色素 c 诱导的凋亡细胞 DNA电泳图0h1h3h4hCKM2h(四)诱导细胞凋亡的因子◆物理性因子:
包括射线(紫外线,? 射线等),较温和的温度刺激(如热激,冷激)等。
◆化学及生物因子:
包括活性氧基团和分子,DNA 和蛋白质合成的抑制剂,激素,细胞生长因子,肿瘤坏死因子 ?(TNF?),抗 Fas/Apo-1/CD95抗体等。
(五)细胞凋亡的检测◆形态学观测:染色法、透射和扫描电镜观察。
◆DNA电泳:DNA片段呈现出梯状条带。
◆TUNEL测定法:即DNA断裂的原位末端标记法。
TUNEL是指末端脱氧核苷酸转移酶介导的dUTP缺口末端标记测定法。此法
可对凋亡细胞的核DNA中产生的3’-OH断裂缺口进行原位标记。
◆彗星电泳法(comet assay):又叫单细胞电泳技术和微凝胶电泳。
用以检测单个细胞中的DNA断裂。细胞凋亡中 DNA ladder 的出现被认为是细胞群发生凋亡的主要标志。细胞在凋亡过程中会有不同的DNA片段化程度,在电场中泳动速度较快,呈现出一种彗星式的图案。正常的无DNA断裂,在电场中泳动速度较慢,呈圆球形。
◆流式细胞分析:细胞凋亡中 DNA断裂,用碘化丙锭使DNA产生激发荧光。IBRVSBV●在细胞凋亡一词出现之前,胚胎学家已观察到动物发育过程中存在着细胞程序性死亡(programmed cell death,PCD)现象,它是胚胎正常发育所必需的。近年来PCD和细胞凋亡常被做为同义词使用,但两者实质上是有差异的。
----首先,PCD是一个功能性概念,描述在一个多细胞生物体中,某些细胞的死亡是个体发育中一个预定的,并受到严格控制的正常组成部分,而凋亡是一个形态学概念,指与细胞坏死不同的受到基因控制的细胞死亡形式;
-----其次,PCD的最终结果是细胞凋亡,但细胞凋亡并非都是程序化的。
2002年10月7日英国人悉尼·布雷诺尔、美国人罗伯特·霍维茨和英国人约翰·苏尔斯顿,因在细胞程序性死亡方面的研究获诺贝尔诺贝尔生理与医学奖。Sydney BrennerH. Robert HorvitzJohn E. Sulston 三、细胞凋亡的分子调控机理细胞凋亡和细胞增殖都是生命的基本现象,是维持体内细胞数量动态平衡的基本措施。在胚胎发育阶段通过细胞凋亡清除多余的和已完成使命的细胞,保证了胚胎的正常发育;在成年阶段通过细胞凋亡清除衰老和病变的细胞,保证了机体的健康。和细胞增殖一样细胞凋亡也是受基因调控的精确过程。
细胞凋亡的途径主要有两条:
★一条是通过胞外信号激活细胞内的凋亡酶 caspase。
★一条是通过线粒体释放凋亡酶激活因子激活 caspase。这些活化
的caspase可将细胞内的重要蛋白降解,引起细胞凋亡。(一)Caspase 家族与凋亡1、Caspase家族
Caspase属于半胱氨酸蛋白酶,相当于线虫中的ced-3,这些蛋白酶是引起细胞凋亡的关键酶,一旦被信号途径激活,能将细胞内的蛋白质降解,使细胞不可逆的走向死亡。它们均有以下特点:
①酶活性依赖于半胱氨酸残基的亲核性;
②裂解靶蛋白位点是天冬氨酸残基后的肽键,所以命名为caspase( Cysteine aspartic acic specific protease ),方便起见称之为凋亡酶;
③都是由两大、两小亚基组成的异四聚体,大、小亚基由同一基因编码,前体被切割后产生两个活性亚基。最早发现人类中与线虫ced-3同源的基因是 ICE (Interleukin-1-converting enzyme) ,即:白介素-1 β转换酶基因,因该酶能将白介素前体切割为活性分子,故名。在人类细胞中已发现11个 ICE 同源物,分为2个亚族:
---ICE 亚族:参与炎症反应
---CED-3 家族,参与细胞凋亡,又分为两类:
★启动者(initiator):如 caspase-8、9,受到信号后,能通过自剪接而
激活,然后引起 caspase 级联反应,如 caspase-8 可依次激活 caspase-3、
6、7。
★执行者(executioner或effector):如caspase-3、6、7,它们可直接降解胞
内的结构蛋白和功能蛋白,引起凋亡,但不能通过自催化或自剪接的方
式激活;
细胞中还具有 caspase 的抑制因子,称为 IAPs(inhibitors of apoptosis
proteins),属于一个庞大的蛋白家族。它们能通过 BIR 结构域(baculovirus IAP repeats domain)与 caspase 结合,抑制其活性,如XIAP。ICE家族成员 A:3类caspase:蓝色参与炎症反应,红色为执行者,绿色为启动者;B:caspase-3的结构模型;C:caspase-3的活化过程执行者启动者参与炎症反应caspase 超家族成员及其相应底物2、Caspase 活化 Caspase 自身以非活化的 Procaspase存在,其激活依赖于其他的 Caspase 在它的天冬氨酸位点裂解活化或自身活化。
Caspase-8自剪切活化,激活 Caspase-3,Caspase-7成为凋亡的执行者。
Caspase-3 激活 Caspase-6。
Caspase 可降解结构蛋白、信号蛋白、转录调控蛋白、周期蛋白等等。
Caspase 还可降解CAD的调节蛋白,释放出CAD,CAD进入细胞核降
解 DNA。
CAD 为caspase-activated Dnase(脱氧核苷酸酶),存在于胞质中。
线虫(C.elegans)凋亡研究发现ced3,ced4基因促进细胞凋亡,ced9基因阻止ced3/ced4的激活,抑制细胞凋亡。Ced3哺乳类同源物是ICE,即Caspase1。
ICE蛋白酶参与 Fas 诱导的细胞凋亡,与线虫细胞凋亡基因 ced-3 同源。
ced-4 在哺乳动物中的同源体为Apaf-1 (apoptotic protease activating factor-1),能激活Caspase3 ,这一过程需要细胞色素c(Apaf-2)和Apaf-3的参与。(二)fas ( CD95)与细胞凋亡 fas 又称作 APO-1,属 TNF 受体和 NGF 受体家族。
1993 年人白细胞分型国际会议统一命名为 CD95。
fas 基因编码产物为分子量 45KD 的跨膜蛋白。
Fas 蛋白与 Fas 配体组成 Fas 系统,二者的结合导致
靶细胞走向凋亡。Fas 具有三个富含半胱氨酸的胞外区和一个称为死亡结构域(Death domain,DD)的胞内区。
Fas 的配体 FasL(Fas ligand)与 Fas 结合后,Fas 三聚化使胞内的DD区构象改变,然后与接头蛋白FADD(Fas-associated death domain)的DD区结合,而后 FADD 的 N 端 DED区(death effector domain)就能与Caspase-8(或-10)前体蛋白结合,形成DISC (death-inducing signaling complex ),引起caspase-8、10通过自身剪激活,它们启动 caspase 的级联反应,使caspase-3、-6、-7激活,这几种Caspase可降解胞内结构蛋白和功能蛋白,最终导致细胞凋亡。=TNF(1) Extrinsic pathway: Fas Signaling PathwayFas (also called Apo-1 or CD95) is a member of the tumor necrosis factor receptor (TNFR) superfamily. Receptor-mediated pathway of apoptosisBcl-2 Family(cytoplasmic factors):
Bad,Bid,and bax:
promote apoptosis;
Bcl-X, Bcl-w,and Bcl-2:
prevent apoptosis.
Internal stimuli:
DNA damage, high Ca2+ ,
Oxidative stress线粒体在 CD95 诱导的凋亡中的作用:
Caspase 引起线粒体 PT(Permeability transition)孔打开,导致线粒体的跨膜电位下降;
细胞色素 C 被释放到胞质中、激活 Caspase-9,后者再激活 Caspase-3。
Bcl-2,Bcl-xl 均作用于线粒体。细胞色素释放引起的凋亡胞外信号分子诱导的细胞凋亡途径 当细胞接受凋亡信号分子(Fas,TNF等)后,凋亡细胞表面信号分子受体相互聚集并与细胞内的衔接蛋白(Adaptor protein)结合,这些衔接蛋白又募集 Procaspases 聚集在受体部, Procaspase 相互活化并产生级联反应,使细胞凋亡。
下游 Caspases 活化后,作用于底物,裂解核纤层蛋白,导致细胞核形成凋亡小体;裂解 Dnase (脱氧核苷酸酶)结合蛋白,使 Dnase 释放,降解 DNA 形成DNA Ladder;裂解参与细胞连接或附着的骨架和其他蛋白,使凋亡细胞皱缩、脱落,便于细胞吞噬;导致膜脂 PS 重排,便于吞噬细胞识别并吞噬。
Procaspases are activated by binding to adaptor proteinsThe caspase cascade involved in apoptosis
Procaspase activation by proteolytic cleavage.
Caspase cascade(2) Intrinsic pathway: Mitochondrial pathwayThe mitochondria-mediated pathway of apoptosisVarious types ofcellular stressBcl-2 family: Bad or Bax to become inserted into OM of MitRelease of cytochrome c from I-O space of Mit.Form a multisubunit complex; and Caspase Cascade,…(三)Bcl2 家族、线粒体与细胞凋亡◆Bcl-2是一种原癌基因,是 ced-9 在哺乳类中的同源物,能抑制细胞凋亡;与线粒体及内质网膜相结合;Bcl-2名称来源于B细胞淋巴瘤/白血病-2(B-cell lymphoma/Leukemia-2,bcl-2)。
◆ bcl-2?蛋白,是膜的整合蛋白,主要存在于线粒体外膜、核膜及部分内质网中。
◆ Bcl-2家族成员都含有1-4个Bcl-2同源结构域(BH1-4),并且通常羧基末端有一穿膜的结构域 (transmembrane region,TM)。其中BH4是抗凋亡蛋白所特有的结构域,BH3是与促进凋亡有关的结构域。
◆根据功能和结构可将Bcl-2基因家族分为两类:
-----抗凋亡的(anti-apoptotic),如:Bcl-2、Bcl-xl、Bcl-w、Mcl-1;
-----促进凋亡的(pro-apoptotic),如:Bax、Bak、Bad、Bid、Bim,在促凋亡蛋白中还有一类仅含BH3结构,如Bid、Bad。BCL-2家族成员Bcl-2家族 引自Katja C. Zimmermann等2001 Bcl-2、线粒体与细胞凋亡◆当 Caspase8 活化后,它一方面作用 Procaspase3,另一方面使Bid 裂解成 2 个片段,其中含 BH3 结构域的 C-端片段被运送到线粒体,与 Bcl-2/Bax 的 BH3 结构域形成复合物,导致细胞色素 C 释放。Cytc 与胞质中Ced4 同源物 Apaf-1(凋亡蛋白酶活化因子apoptosis protease activating factor)结合并活Apaf-1,活化的 Apaf-1 再活化 Procaspase9,最后引起细胞凋亡。
Molecular pathways of apoptosisActivation of caspase-2 is required for permeabilization of mitochondria, release of cytochrome c, and apoptosisPathways to cell death in C. elegans and mammals(四)p53与细胞凋亡p53 是一种抑癌基因,其生物学功能是在G期监视DNA的完整性。如有损伤,则抑制细胞增殖,直到DNA修复完成。如果DNA不能被修复,则诱导其调亡,
在依赖P53蛋白的细胞凋亡中,P53蛋白能特异地抑制 Bcl2 的表达,但对 Bax 的表达则有明显的促进作用。在这些细胞中, P53蛋白的积累和活动引起了细胞凋亡。
(五)细胞凋亡的信号转导Ras-PI3-K-Akt(PKB)抗细胞凋亡途径:
----Ras-PI3-K-Akt(PKB)-Caspase9磷酸化,使其不能参与Caspase3
的活化。
----Ras-PI3-K-Akt(PKB)-Bad磷酸化,使其失去阻断Bcl2的功能。
NF-kB与细胞凋亡:
NF-kB是一种调节基因转录的核因子。 NF-kB的活化抑制了Caspase8的活化,从而阻值了下游Caspase活化以及细胞凋亡。
ASK1-SAPK/JNK及ASK1-p38信号途径:
----ASK1(apoptosis signal regulating kinase),是一种哺乳类的MAPKKK。在哺乳类细胞中,有三套MAPK信号组件:RaF-MAPKK-MAPKK,MEKK-MKK4-JNK,MKK3-p38(一种MAPK)。
----ASK1能选择地活化MEKK-MKK4-JNK,MKK3-p38(一种MAPK),引起细胞凋亡。机体结构细胞增殖细胞分化细胞凋亡细胞信号转导染色体(DNA与蛋白质的相互作用)衰老全教程到此结束,谢谢大家!
第二章 细胞基本知识概要
第三章 细胞生物学研究方法
第四章 细胞质膜与细胞表面
第五章 物质跨膜运输与信号传递
第六章 细胞质基质与内膜系统
第七章 细胞的能量转换
第八章 细胞核与染色体
第九章 核糖体
第 十 章 细胞骨架
第十一章 细胞增殖及其调控
第十二章 细胞分化与基因表达调控
第十三章 细胞衰老与凋亡
第十三章 细胞衰老和凋亡第一节 细胞衰老(cellular aging或cell senescence)又称老化,通常指生物发育成熟后,在正常情况下随着年龄的增加,机能减退,内环境稳定性下降,结构中心组分退行性变化,趋向死亡的不可逆的现象。
衰老和死亡是生命的基本现象,衰老过程发生在生物界的整体水平、种群水平、个体水平、细胞水平以及分子水平等不同的层次。一、Hayflick 界限(Hayflick Limitation)● Hayflick 界限:细胞,至少是培养的细胞,不是不死的,而是有一定的寿命;它们的增殖能力不是无限的,而是有一定的界限。“不死性”学说的错误在于:
◆癌细胞或培养的细胞系是不正常细胞,其染色体数目或形态已经不同于原先的细胞。
◆细胞的增殖能力与供体年龄有关。
◆物种寿命与培养细胞寿命之间存在着一定的关系。
◆衰老的原因在于细胞本身。不同年龄来源的人成纤维细胞的增殖代数二、细胞在体内条件下的衰老●在机体内,细胞的衰老和死亡是常见的现象,甚至在个体发育的早期也会发生。
●正常情况下终生保持分裂的细胞,其分裂能力是否随着有机体年龄的增高而下降?它们 会不会衰老?
◆衰老动物体内,细胞分裂速度显著减慢,其原因主要是G1期明
显延长。
◆衰老个体内的环境因素影响了细胞的增殖和衰老。
◆骨髓干细胞移植实验说明随着年龄的增加,干细胞增殖速度也
趋缓慢。三、衰老细胞结构的变化●细胞核的变化:核膜内折,染色质固缩化。
●内质网的变化: 衰老动物内质网成分弥散性地分散于核周胞质中,粗面内质网的总量似乎是减少了。
●线粒体的变化:通常,细胞中线粒体的数量随年龄增大而减少,而其体积则随年龄增大而增大。
●致密体的生成:由溶酶体和线粒体转化而来。
●膜系统的变化
◆衰老的细胞,其膜流动性降低、韧性减小。
◆衰老细胞间间隙连接及膜内颗粒的分布也发生变化。(一)形态变化核 增大、染色深、核内有包含物
染色质 凝聚、固缩、碎裂、溶解
质膜 粘度增加、流动性降低
细胞质 色素积聚、空泡形成
线粒体 数目减少、体积增大、mt DNA突变或丢失
高尔基体 碎裂
尼氏体 消失
包含物 糖原减少、脂肪积聚
核膜 内陷表15-1 衰老细胞的形态变化尼氏体:围绕某些感觉神经末梢的卵圆形同心层状结构 。1、DNA:复制与转录受阻,端粒DNA、mtDNA缺失。DNA氧化、断裂、缺失和交联,甲基化程度降低。
2、RNA:m RNA和t RNA含量降低。
3、蛋白质:含成下降,细胞内蛋白质发生糖基化、氨甲酰化、脱氨基等修饰反应,导致蛋白质稳定性、抗原性,可消化性下降,自由基使蛋白质肽断裂,交联而变性。氨基酸由左旋变为右旋。
4、酶分子:活性中心被氧化,金属离子丢失,酶分子的二级结构,溶解度,等电点发生改变,酶失活。
5、脂类:不饱和脂肪酸被氧化。引起膜脂之间或与脂蛋白之间交联,膜的流动性降低。 (二)分子水平的变化四、细胞衰老的分子机理 该理论认为,代谢过程中产生的活性氧基团或分子(reactive oxygen species,ROS)引发的氧化性损伤的积累,最终导致衰老。
自由基:是一类瞬时形成的含不成对电子的原子或功能基团,普遍存在于生物系统。具有高度反应活性,可引发链式自由基反应,引起DNA、蛋白质和脂类,尤其是多不饱和脂肪酸等大分子物质变性和交联,损伤DNA、生物膜、重要的结构蛋白和功能蛋白,从而引起衰老各种现象的发生。
ROS主要有三种类型:
① O2-,即超氧自由基; ② OH-,即羟自由基; ③ H2O2。
(一)氧化性损伤学说:内源性自由基是人体自由基的主要来源,其产生的主要途径有:
①由线粒体呼吸链电子泄漏产生;
②由经过氧化物酶体的多功能氧化酶(MFO)等催化底物羟化产生。
③机体血红蛋白、肌红蛋白中还可通过非酶促反应产生自由基。
清除ROS,就可以延长寿命。正常细胞内存在清除自由基的防御系统,包括酶系统和非酶系统:
酶系统:SOD,CAT,GR等;
非酶系统:维生素E,醌类等电子受体。
(二)端粒与衰老: 细胞增殖次数与端粒DNA长度有关。
体细胞染色体的端粒DNA会随细胞分裂次数增加而不断缩短。细胞DNA每复制一次端粒就缩短一段,当缩短到一定程度至Hayflick点时,可能会启动DNA损伤检测点(DNA damage checkpoint),激活p53,引起p21表达,导致不可逆地退出细胞周期,走向衰亡。资料表明人的成纤维细胞端粒每年缩短14-18bp,可见染色体的端粒有细胞分裂计数器的功能,能记忆细胞分裂的次数。
端粒的长度与端粒酶(telomerase)的活性有关。
端粒酶是一种反转录酶,能以自身的RNA为模板合成端粒DNA,在精原细胞、干细胞和肿瘤细胞(如Hela细胞)中有较高的端粒酶活性,而正常体细胞中端粒酶的活性很低,呈抑制状态。(三)r DNA与衰老
在S.cerevisiae该酵母细胞中,会通过同源重组形成染色体外rDNA环
(extrachromosomal rDNA circle, ESC)。当ESC积累到500~1000拷贝是,细胞就进入衰老。但在哺乳动物中ESC并不多见。
(四)沉默信息调节蛋白(Sir )复合物与衰老
Sir复合物的功能是阻值它们所在位点的 DNA 转录。
(五)SGS1基因、WRN基因与衰老
SGS1基因编码的蛋白属于RecQ亚族的DNA解旋酶。WRN基因与SGS1基因同源,属于DNA解旋酶。在Werner综合症(WS)的早衰患者中观察不到WRN蛋白。Werner's syndrome 成人早衰症(Werner's syndrome)病人平均39岁时出现衰老,47岁左右生命结束,患婴幼儿早衰症(Hutchinson-Gilford syndrome,图15-2)的小孩在1岁时出现明显的衰老,12-18岁即过早夭折。
(六)程序性衰老(programmed senescence) 程序性衰老理论认为:生物的生长、发育、衰老和死亡都由基因程序控制的,衰老实际上是某些基因依次开启或关闭的结果。
例如在小鼠肝中,胚胎早期表达的胞质丙氨酸转氨酶(cytosolic alanine aminotransferase,c AAT)为 A 型,随后停止表达,但是在衰老时则表达 B 型 c AAT,其它类似的衰老标志物(senescence markers)也有报道,如肝脏中的衰老标志蛋白 2(senescence marker protein 2)也是在老年期表达。
(七)线粒体 DNA 突变(mitochondrial DNA mutation) 在线粒体氧化磷酸化生成 ATP 的过程中,大约有1-4%氧转化ROS,因此线粒体是自由基浓度最高的细胞器。 mtDNA裸露于基质,缺乏结合蛋白的保护,最易受自由基伤害,而催化mtDNA复制的DNA聚合酶γ不具有校正功能,复制错误频率高,同时缺乏有效的修复酶,故mtDNA最容易发生突变。Mt DNA突变使呼吸链功能受损,进一步引起自由基堆积,如此反复循环。衰老个体细胞中mtDNA缺失表现明显,并随着年龄的增加而增加,许多研究认为 mtDNA缺失与衰老及伴随的老年衰退性疾病有密切关系。
人类的脑、心、骨骼肌的氧负荷(oxidative stress)最大,因而是最容易衰老的组织。研究表明,限制热量(caloric restriction)摄入能明显延长小鼠的寿命。第二节 细胞凋亡 (Apoptosis)一、细胞凋亡的概念及其生物学意义● 概念:
细胞凋亡是一个主动的由基因决定的自动结束生命的过程,也常常被称为细胞程序死亡(programmed cell death, PCD)。凋亡细胞将被吞噬细胞吞噬。
● 细胞调亡的生物学意义:
它是维持组织机能和形态所必需的。生物发育过程中及成体组织中正常的细胞凋亡有助于保证细胞只在需要它们的时候和需要它们活的地方存活。这对于多细胞生物个体发育的正常进行,自稳平衡的保持以及抵御外界各种因素 的干扰方面都起着非常关键的作用。 蝌蚪尾的消失
发育过程中手和足
的成形过程 脊椎动物的神经系统的发育 脊椎动物的神经
系统的发育 二、细胞凋亡的形态学和生物化学特征 细胞死亡的方式通常有3种:即①细胞坏死(necrosis)。②细胞凋亡(apoptosis)。③细胞程序性死亡(programmed cell death,PCD)。
(一)细胞坏死
是细胞受到化学因素(如强酸、强碱、有毒物质)、物理因素(如热、辐射)和生物因素(如病原体)等环境因素的伤害,引起细胞死亡的现象。
早期表现为胞质内线粒体和内质网肿胀、崩解,核发生固缩或断裂。随着胞质内蛋白变性、凝固或碎裂,以及嗜碱性核蛋白的降解,细胞质呈现强嗜酸性,故坏死组织或细胞在苏木精/伊红染色切片中,胞质呈均一的深伊红色,原有的微细结构消失。
在含水量高的细胞,可因胞质内水泡不断增大,并发生溶解,导致细胞结构完全消失,最后细胞膜和细胞器破裂,DNA降解,细胞内容物流出,引起周围组织炎症反应。(二)细胞凋亡的形态学特征Kerr( 1972)最先提出,与细胞坏死的区别是:
①染色质聚集、分块、位于核膜上,胞质凝缩,最后核断裂,细胞通过出芽的方式形成许多凋亡小体(apoptotic bodies);
②凋亡小体内有结构完整的细胞器,还有凝缩的染色体,可被邻近细胞吞噬消化,因始终有膜封闭,没有内溶物释放,故不会引起炎症;
③凋亡细胞中仍需要合成一些蛋白质,但是在坏死细胞中ATP和蛋白质合成受阻或终止;
④核酸内切酶活化,导致染色质 DNA 在核小体连接部位断裂,形成约 200bp 整数倍的核酸片段,凝胶电泳图谱呈梯状;
⑤凋亡通常是生理性变化,而细胞坏死是病理性变化。
表15-2 细胞凋亡和细胞坏死的区别区别点 细胞凋亡 细胞坏死
起因 生理或病理性 病理性变化或剧烈损伤
范围 单个散在细胞 大片组织或成群细胞
细胞膜 保持完整,一直到形成凋亡小体 破损
染色质 凝聚在核膜下呈半月状 呈絮状
细胞器 无明显变化 肿胀、内质网崩解
细胞体积 固缩变小 肿胀变大
凋亡小体 有,被邻近细胞或 无,细胞自溶,残余碎片
巨噬细胞吞噬 被巨噬细胞吞噬
基因组DNA 有控降解,电泳图谱呈梯状 随机降解,电泳图谱呈涂抹状
蛋白质合成 有 无
调节过程 受基因调控 被动进行
炎症反应 无,不释放细胞内容物 有,释放内容物。细胞坏死与凋亡的形态区别坏死细胞凋亡细胞细胞凋亡的形态学特征? Morphologi changes:
Early : Chromosome condensation, cell body shrink
Later : Blebbing and Nucleus and cytoplasm fragment—Apoptotic bodies
At last: Phagocytosed弯曲胸腺细胞正常凋亡正常凋亡(三)细胞凋亡的生化特征核酸内切酶活化,导致染色质DNA在核小体连接部位断裂,形成约200bp整数倍的核酸片段,凝胶电泳图谱呈梯状
细胞色素 c 诱导的凋亡细胞 DNA电泳图0h1h3h4hCKM2h(四)诱导细胞凋亡的因子◆物理性因子:
包括射线(紫外线,? 射线等),较温和的温度刺激(如热激,冷激)等。
◆化学及生物因子:
包括活性氧基团和分子,DNA 和蛋白质合成的抑制剂,激素,细胞生长因子,肿瘤坏死因子 ?(TNF?),抗 Fas/Apo-1/CD95抗体等。
(五)细胞凋亡的检测◆形态学观测:染色法、透射和扫描电镜观察。
◆DNA电泳:DNA片段呈现出梯状条带。
◆TUNEL测定法:即DNA断裂的原位末端标记法。
TUNEL是指末端脱氧核苷酸转移酶介导的dUTP缺口末端标记测定法。此法
可对凋亡细胞的核DNA中产生的3’-OH断裂缺口进行原位标记。
◆彗星电泳法(comet assay):又叫单细胞电泳技术和微凝胶电泳。
用以检测单个细胞中的DNA断裂。细胞凋亡中 DNA ladder 的出现被认为是细胞群发生凋亡的主要标志。细胞在凋亡过程中会有不同的DNA片段化程度,在电场中泳动速度较快,呈现出一种彗星式的图案。正常的无DNA断裂,在电场中泳动速度较慢,呈圆球形。
◆流式细胞分析:细胞凋亡中 DNA断裂,用碘化丙锭使DNA产生激发荧光。IBRVSBV●在细胞凋亡一词出现之前,胚胎学家已观察到动物发育过程中存在着细胞程序性死亡(programmed cell death,PCD)现象,它是胚胎正常发育所必需的。近年来PCD和细胞凋亡常被做为同义词使用,但两者实质上是有差异的。
----首先,PCD是一个功能性概念,描述在一个多细胞生物体中,某些细胞的死亡是个体发育中一个预定的,并受到严格控制的正常组成部分,而凋亡是一个形态学概念,指与细胞坏死不同的受到基因控制的细胞死亡形式;
-----其次,PCD的最终结果是细胞凋亡,但细胞凋亡并非都是程序化的。
2002年10月7日英国人悉尼·布雷诺尔、美国人罗伯特·霍维茨和英国人约翰·苏尔斯顿,因在细胞程序性死亡方面的研究获诺贝尔诺贝尔生理与医学奖。Sydney BrennerH. Robert HorvitzJohn E. Sulston 三、细胞凋亡的分子调控机理细胞凋亡和细胞增殖都是生命的基本现象,是维持体内细胞数量动态平衡的基本措施。在胚胎发育阶段通过细胞凋亡清除多余的和已完成使命的细胞,保证了胚胎的正常发育;在成年阶段通过细胞凋亡清除衰老和病变的细胞,保证了机体的健康。和细胞增殖一样细胞凋亡也是受基因调控的精确过程。
细胞凋亡的途径主要有两条:
★一条是通过胞外信号激活细胞内的凋亡酶 caspase。
★一条是通过线粒体释放凋亡酶激活因子激活 caspase。这些活化
的caspase可将细胞内的重要蛋白降解,引起细胞凋亡。(一)Caspase 家族与凋亡1、Caspase家族
Caspase属于半胱氨酸蛋白酶,相当于线虫中的ced-3,这些蛋白酶是引起细胞凋亡的关键酶,一旦被信号途径激活,能将细胞内的蛋白质降解,使细胞不可逆的走向死亡。它们均有以下特点:
①酶活性依赖于半胱氨酸残基的亲核性;
②裂解靶蛋白位点是天冬氨酸残基后的肽键,所以命名为caspase( Cysteine aspartic acic specific protease ),方便起见称之为凋亡酶;
③都是由两大、两小亚基组成的异四聚体,大、小亚基由同一基因编码,前体被切割后产生两个活性亚基。最早发现人类中与线虫ced-3同源的基因是 ICE (Interleukin-1-converting enzyme) ,即:白介素-1 β转换酶基因,因该酶能将白介素前体切割为活性分子,故名。在人类细胞中已发现11个 ICE 同源物,分为2个亚族:
---ICE 亚族:参与炎症反应
---CED-3 家族,参与细胞凋亡,又分为两类:
★启动者(initiator):如 caspase-8、9,受到信号后,能通过自剪接而
激活,然后引起 caspase 级联反应,如 caspase-8 可依次激活 caspase-3、
6、7。
★执行者(executioner或effector):如caspase-3、6、7,它们可直接降解胞
内的结构蛋白和功能蛋白,引起凋亡,但不能通过自催化或自剪接的方
式激活;
细胞中还具有 caspase 的抑制因子,称为 IAPs(inhibitors of apoptosis
proteins),属于一个庞大的蛋白家族。它们能通过 BIR 结构域(baculovirus IAP repeats domain)与 caspase 结合,抑制其活性,如XIAP。ICE家族成员 A:3类caspase:蓝色参与炎症反应,红色为执行者,绿色为启动者;B:caspase-3的结构模型;C:caspase-3的活化过程执行者启动者参与炎症反应caspase 超家族成员及其相应底物2、Caspase 活化 Caspase 自身以非活化的 Procaspase存在,其激活依赖于其他的 Caspase 在它的天冬氨酸位点裂解活化或自身活化。
Caspase-8自剪切活化,激活 Caspase-3,Caspase-7成为凋亡的执行者。
Caspase-3 激活 Caspase-6。
Caspase 可降解结构蛋白、信号蛋白、转录调控蛋白、周期蛋白等等。
Caspase 还可降解CAD的调节蛋白,释放出CAD,CAD进入细胞核降
解 DNA。
CAD 为caspase-activated Dnase(脱氧核苷酸酶),存在于胞质中。
线虫(C.elegans)凋亡研究发现ced3,ced4基因促进细胞凋亡,ced9基因阻止ced3/ced4的激活,抑制细胞凋亡。Ced3哺乳类同源物是ICE,即Caspase1。
ICE蛋白酶参与 Fas 诱导的细胞凋亡,与线虫细胞凋亡基因 ced-3 同源。
ced-4 在哺乳动物中的同源体为Apaf-1 (apoptotic protease activating factor-1),能激活Caspase3 ,这一过程需要细胞色素c(Apaf-2)和Apaf-3的参与。(二)fas ( CD95)与细胞凋亡 fas 又称作 APO-1,属 TNF 受体和 NGF 受体家族。
1993 年人白细胞分型国际会议统一命名为 CD95。
fas 基因编码产物为分子量 45KD 的跨膜蛋白。
Fas 蛋白与 Fas 配体组成 Fas 系统,二者的结合导致
靶细胞走向凋亡。Fas 具有三个富含半胱氨酸的胞外区和一个称为死亡结构域(Death domain,DD)的胞内区。
Fas 的配体 FasL(Fas ligand)与 Fas 结合后,Fas 三聚化使胞内的DD区构象改变,然后与接头蛋白FADD(Fas-associated death domain)的DD区结合,而后 FADD 的 N 端 DED区(death effector domain)就能与Caspase-8(或-10)前体蛋白结合,形成DISC (death-inducing signaling complex ),引起caspase-8、10通过自身剪激活,它们启动 caspase 的级联反应,使caspase-3、-6、-7激活,这几种Caspase可降解胞内结构蛋白和功能蛋白,最终导致细胞凋亡。=TNF(1) Extrinsic pathway: Fas Signaling PathwayFas (also called Apo-1 or CD95) is a member of the tumor necrosis factor receptor (TNFR) superfamily. Receptor-mediated pathway of apoptosisBcl-2 Family(cytoplasmic factors):
Bad,Bid,and bax:
promote apoptosis;
Bcl-X, Bcl-w,and Bcl-2:
prevent apoptosis.
Internal stimuli:
DNA damage, high Ca2+ ,
Oxidative stress线粒体在 CD95 诱导的凋亡中的作用:
Caspase 引起线粒体 PT(Permeability transition)孔打开,导致线粒体的跨膜电位下降;
细胞色素 C 被释放到胞质中、激活 Caspase-9,后者再激活 Caspase-3。
Bcl-2,Bcl-xl 均作用于线粒体。细胞色素释放引起的凋亡胞外信号分子诱导的细胞凋亡途径 当细胞接受凋亡信号分子(Fas,TNF等)后,凋亡细胞表面信号分子受体相互聚集并与细胞内的衔接蛋白(Adaptor protein)结合,这些衔接蛋白又募集 Procaspases 聚集在受体部, Procaspase 相互活化并产生级联反应,使细胞凋亡。
下游 Caspases 活化后,作用于底物,裂解核纤层蛋白,导致细胞核形成凋亡小体;裂解 Dnase (脱氧核苷酸酶)结合蛋白,使 Dnase 释放,降解 DNA 形成DNA Ladder;裂解参与细胞连接或附着的骨架和其他蛋白,使凋亡细胞皱缩、脱落,便于细胞吞噬;导致膜脂 PS 重排,便于吞噬细胞识别并吞噬。
Procaspases are activated by binding to adaptor proteinsThe caspase cascade involved in apoptosis
Procaspase activation by proteolytic cleavage.
Caspase cascade(2) Intrinsic pathway: Mitochondrial pathwayThe mitochondria-mediated pathway of apoptosisVarious types ofcellular stressBcl-2 family: Bad or Bax to become inserted into OM of MitRelease of cytochrome c from I-O space of Mit.Form a multisubunit complex; and Caspase Cascade,…(三)Bcl2 家族、线粒体与细胞凋亡◆Bcl-2是一种原癌基因,是 ced-9 在哺乳类中的同源物,能抑制细胞凋亡;与线粒体及内质网膜相结合;Bcl-2名称来源于B细胞淋巴瘤/白血病-2(B-cell lymphoma/Leukemia-2,bcl-2)。
◆ bcl-2?蛋白,是膜的整合蛋白,主要存在于线粒体外膜、核膜及部分内质网中。
◆ Bcl-2家族成员都含有1-4个Bcl-2同源结构域(BH1-4),并且通常羧基末端有一穿膜的结构域 (transmembrane region,TM)。其中BH4是抗凋亡蛋白所特有的结构域,BH3是与促进凋亡有关的结构域。
◆根据功能和结构可将Bcl-2基因家族分为两类:
-----抗凋亡的(anti-apoptotic),如:Bcl-2、Bcl-xl、Bcl-w、Mcl-1;
-----促进凋亡的(pro-apoptotic),如:Bax、Bak、Bad、Bid、Bim,在促凋亡蛋白中还有一类仅含BH3结构,如Bid、Bad。BCL-2家族成员Bcl-2家族 引自Katja C. Zimmermann等2001 Bcl-2、线粒体与细胞凋亡◆当 Caspase8 活化后,它一方面作用 Procaspase3,另一方面使Bid 裂解成 2 个片段,其中含 BH3 结构域的 C-端片段被运送到线粒体,与 Bcl-2/Bax 的 BH3 结构域形成复合物,导致细胞色素 C 释放。Cytc 与胞质中Ced4 同源物 Apaf-1(凋亡蛋白酶活化因子apoptosis protease activating factor)结合并活Apaf-1,活化的 Apaf-1 再活化 Procaspase9,最后引起细胞凋亡。
Molecular pathways of apoptosisActivation of caspase-2 is required for permeabilization of mitochondria, release of cytochrome c, and apoptosisPathways to cell death in C. elegans and mammals(四)p53与细胞凋亡p53 是一种抑癌基因,其生物学功能是在G期监视DNA的完整性。如有损伤,则抑制细胞增殖,直到DNA修复完成。如果DNA不能被修复,则诱导其调亡,
在依赖P53蛋白的细胞凋亡中,P53蛋白能特异地抑制 Bcl2 的表达,但对 Bax 的表达则有明显的促进作用。在这些细胞中, P53蛋白的积累和活动引起了细胞凋亡。
(五)细胞凋亡的信号转导Ras-PI3-K-Akt(PKB)抗细胞凋亡途径:
----Ras-PI3-K-Akt(PKB)-Caspase9磷酸化,使其不能参与Caspase3
的活化。
----Ras-PI3-K-Akt(PKB)-Bad磷酸化,使其失去阻断Bcl2的功能。
NF-kB与细胞凋亡:
NF-kB是一种调节基因转录的核因子。 NF-kB的活化抑制了Caspase8的活化,从而阻值了下游Caspase活化以及细胞凋亡。
ASK1-SAPK/JNK及ASK1-p38信号途径:
----ASK1(apoptosis signal regulating kinase),是一种哺乳类的MAPKKK。在哺乳类细胞中,有三套MAPK信号组件:RaF-MAPKK-MAPKK,MEKK-MKK4-JNK,MKK3-p38(一种MAPK)。
----ASK1能选择地活化MEKK-MKK4-JNK,MKK3-p38(一种MAPK),引起细胞凋亡。机体结构细胞增殖细胞分化细胞凋亡细胞信号转导染色体(DNA与蛋白质的相互作用)衰老全教程到此结束,谢谢大家!
同课章节目录
- 第一章 走近细胞
- 第1节 从生物圈到细胞
- 第2节 细胞的多样性和统一性
- 第二章 组成细胞的分子
- 第1节 细胞中的元素和化合物
- 第2节 生命活动的主要承担者──蛋白质
- 第3节 遗传信息的携带者──核酸
- 第4节 细胞中的糖类和脂质
- 第5节 细胞中的无机物
- 第三章 细胞的基本结构
- 第1节 细胞膜──系统的边界
- 第2节 细胞器──系统内的分工合作
- 第3节 细胞核──系统的控制中心
- 第四章 细胞的物质输入和输出
- 第1节 物质跨膜运输的实例
- 第2节 生物膜的流动镶嵌模型
- 第3节 物质跨膜运输的方式
- 第五章 细胞的能量供应和利用
- 第1节 降低化学反应活化能的酶
- 第2节 细胞的能量“通货”──ATP
- 第3节 ATP的主要来源──细胞呼吸
- 第4节 能量之源——光与光合作用
- 第六章 细胞的生命历程
- 第1节 细胞的增殖
- 第2节 细胞的分化
- 第3节 细胞的衰老和凋亡
- 第4节 细胞的癌变