人教版生物必修二第五章第二节染色体变异
文档属性
| 名称 | 人教版生物必修二第五章第二节染色体变异 |

|
|
| 格式 | zip | ||
| 文件大小 | 6.5MB | ||
| 资源类型 | 教案 | ||
| 版本资源 | 人教版(新课程标准) | ||
| 科目 | 生物学 | ||
| 更新时间 | 2014-12-10 00:00:00 | ||
图片预览






文档简介
课件159张PPT。第6章 染色体变异第1节 染色体结构变异
第2节 染色体数目变异
第一节 染色体结构变异结构变异的形成:断裂—重接
◎使染色体产生折断的因素:
自然:温度剧变、营养生理条件异常、遗传因素等;
人为:物理射线与化学药剂处理等。
◎染色体折断的结果:
正确重接:重新愈合,恢复原状;
错误重接:产生结构变异;
保持断头:产生结构变异。
◎结构变异的基本类型:
缺失、重复、倒位、易位
概念、类型与特点;
细胞学特征与鉴定;
遗传效应。第一节 染色体结构变异◎ 缺失
◎ 重复
◎ 倒位
◎ 易位
◎ 染色体结构变异的应用(一)、缺失 的类型
缺失:染色体的某一区段丢失了
断片:缺失的区段无着丝粒
顶端缺失:缺失的区段为某臂的外端
某一整臂缺失了就成为顶端着丝点染色体
中间缺失:缺失的区段为某臂的内段 顶端缺失染色体很难定型,常会重接。
(1)断头很难愈合,断头可能同另一有着丝粒的染色体的断头重接,成为单着粒染色体
(2)顶端缺失染色体的两个姊妹染色单体可能在断头上彼此接合,形成双着丝粒染色体
双着丝粒染色体就会在细胞分裂的后期受两个着丝粒向相反两极移动所产生的拉力所折断,再次造成结构的变异而不能稳定,形成断裂—融合—桥 断裂—融合—桥复制
姊妹染色单体顶端断头连接(融合)
有丝分裂后期桥(桥)
新的断裂
(二)、缺失的细胞学鉴定无着丝粒断片;
最初发生缺失的细胞在分裂时可见无着丝粒断片。
缺失环(环形或瘤形突出);
中间缺失杂合体偶线期和粗线期出现;
二价体末端突出;
顶端缺失杂合体粗线期、双线期,交叉未完全端化的二价体末端不等长。缺失的细胞学特征果蝇唾腺染色体的缺失圈(三)、缺失的遗传效应缺失纯合体:
致死或半致死。
缺失杂合体:
缺失区段较长时,生活力差、配子(尤其是花粉)败育或竞争不过正常配子;
缺失区段较小时,可能会造成假显性现象或其它异常现象(p117例:猫叫综合症)。缺失区段上基因丢失导致:
基因所决定、控制的生物功能丧失或异常;
基因间相互作用关系破坏;
基因排列位置关系改变。
影响缺失对生物个体危害程度的因素:
缺失区段的大小;
缺失区段所含基因的多少;
缺失基因的重要程度;
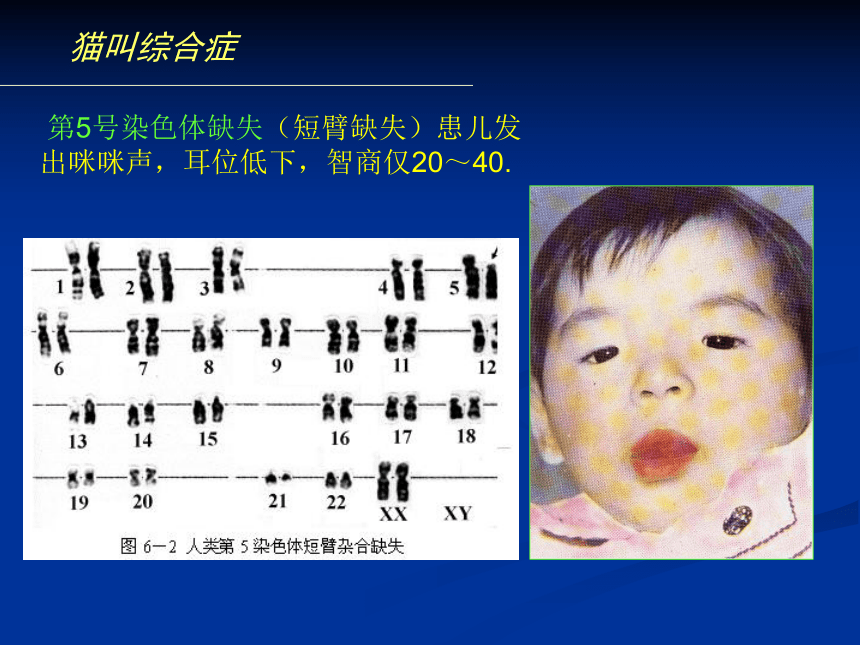
染色体倍性水平。缺失杂合体的假显性现象猫叫综合症 第5号染色体缺失(短臂缺失)患儿发
出咪咪声,耳位低下,智商仅20~40.猫叫综合症“猫叫综合症”是第5号染色体短臂缺失引起的遗传病,发生率为十万分之一,在国内外均很少见。患儿一般表现为生长发育迟缓,头央部畸形,哭声轻,音调高,皮纹改变等特点,并有严重的智能障碍,而其最明显的特征是哭声类似猫叫,“猫叫综合症”因此而得名。据称,病儿哭声异常可能系喉部发育不良所致,也可能与脑损害有关。 二、重复(duplication)(一)、重复的类别与形成
重复:染色体多了自己的某一区段。
顺接重复(tanden duplication) ;
反接重复(reverse duplication) 。(二)、重复的细胞学鉴定重复圈(环)果蝇X染色体上16A区段重复的形成 重复区段内不能有着丝粒,否则重复染色体就变成双着丝粒的染色体,就会继续发生结构变异,很难稳定成型。
重复和缺失总是伴随出现的。某染色体的一个区段转移给同源的另一个染色体之后,它自己就成为缺失染色体了。 (三)、重复的遗传效应重复对个体综合表现的影响:
重复区段内的基因重复,影响基因间的平衡关系;
会影响个体的生活力(影响的程度与重复区段的大小有关)。
剂量效应(dosage effect):
果蝇眼色遗传的剂量效应,红色(V+)对朱红(V)为显性;
杂合体(V+V)表现为红色;
但(V+VV)的表现型却为朱红色。
位置效应(position effect):
果蝇眼面大小遗传的位置效应;
位置效应的意义。果蝇棒眼的位置效应三、倒位(inversion)(一)、倒位的类别与形成
倒位:染色体某区段的正常直线顺序颠倒了
臂内倒位(paracentric inversion);
臂间倒位(pericentic inversion)。(二)、倒位的细胞学鉴定倒位杂合体的联会倒位杂合体的“倒位圈”(三)、倒位的遗传效应1. 倒位杂合体的部分不育现象:
倒位圈内发生交换后,产生的交换型配子(50%)含重复缺失染色单体,这类配子是不育的;
只有部分孢母细胞在减数分裂时倒位圈内会发生非姊妹染色单体间的交换;
倒位点可以当作一个显性基因位点看待,其性状表现就是倒位杂合体部分不育。
2. 倒位改变了基因在染色体上的排列:
基因间距离关系发生改变;
可能引起倒位区段基因的位置效应;
倒位杂合体的基因间交换值降低:
倒位圈的结构影响联会复合体的正常形成;
倒位圈内发生交换后产生的交换型配子是不育的。
3. 是物种进化的重要因素之一。
倒位可能导致新物种的产生。3、倒位的遗传效应 (1)位置效应:倒位区段内、外各个基因之间的物理距离发生改变,其遗传距离一般也改变。 5a倒位杂合体的重组率(Pr-bt1)0.4 %、(bm1-bt1)0%、(a2-bt1)4.5%(2)降低倒位杂合体上连锁基因的重组率:产生的交换型配子数明显减少,故重组率降低。 (3)倒位可以形成新种,促进生物进化例如:果蝇(n=4):不同倒位特点的种,分布在不同地理区域。倒位杂合体的交换臂间倒位杂合体的交换臂内倒位杂合体的交换臂内倒位形成的“后期 I 桥”四、易位(translocation)(一)、易位的类别与形成
相互易位(reciprocal translocation)
简单易位(simple translocation)(转移)
易位的形成(
易位染色体
易位染色体的表示方法
易位杂合体(translocation heterozygote)
易位纯合体(translocation homozygote)1221112212易位的类别与形成(二)、易位的细胞学鉴定易位杂合体的联会和分离易位杂合体的联会和分离1,121221121,122,212,21全部不育配子相邻式分离11122212,122,121,211,21全部不育配子相邻式分离2全部可育配子交替式分离易位杂合体粗线期十字形配对易位杂合体的联会和分离玉米(2n=20)终变期染色体(三)、易位的遗传效应易位杂合体半不育现象
易位杂合体十字形配对进行交替式分离所产生的配子是可育的,而进行相邻式分离产生的配子是不育的。
花粉50%败育,胚囊50%败育,结实率只有50%,由半不育植株的种子所长出的植株又会有半数是半不育的,半数是正常可育的。
易位杂合体后期分离时:
1,21/12,2 相邻式,重复缺失-败育,1/2
1,12/2,21
1,2/12,21 交替式,正常/易位-可育,1/2(三)、易位的遗传效应易位杂合体半不育现象
玉米型:相邻式与交换式各占50%,配子半不育,可视为一个半不育显性基因看待。
月见草型:全部采用交替式分离,所有配子都可育。
曼驼罗、风铃草、紫万年青, 易位杂合体后期分离100%是交替式 。
(三)、易位的遗传效应易位改变了生物的连锁群
基因间连锁关系和位置效应变化
易位杂合体的基因重组值降低
十字形结构影响联会复合体中交换的正常形成
易位与生物进化和新物种形成
相互易位纯合体可以保存形成的新的连锁群关系
易位可能导致染色体融合,引起染色体数目变异染色体融合易位造成染色体融合,导致染色体数目的变异
AB丢失Robertsonian translocation五、 染色体结构变异的应用(一)、利用结构变异进行基因定位
利用缺失的假显性现象将基因定位到染色体某一个区段
以易位点为遗传标记(半不育)进行连锁分析
(二)、果蝇的ClB测定
(三)、利用易位创造玉米不育系的保持系
(四)、易位在家蚕生产上的应用1、利用缺失造成的假显性现象,可以进行基因定位
使载有显性基因的染色体发生缺失,让其隐性等位基因表现“假显性” 对表现假显性的个体进行细胞学鉴定,发现某染色体缺失了某一区段,就说明该显性基因位于该染色体的缺失区段上。 (一)、利用结构变异进行基因定位(一)、利用结构变异进行基因定位
2、以易位点为遗传标记(半不育)进行连锁分析易位杂交体产生的可育配子中一半含两个正常染色体(1,2),一半含两个易位染色体(12,21)。那么在易位杂合体自交子代群体内, 1/4正常可育个体, 2/4半不育易位杂合体, 1/4可育易位纯合体
易位染色体的易位接合点相当于一个半不育的显性遗传基因(T),正常染色体与易位接合点相对的等位点,则相当于一个可育的隐性遗传基因(t)
遗传学上利用这一特点,用两点或三点测验,根据T–t与某邻近基因之间的重组率,确定易位接合点在染色体上的位置 。
例:玉米长节间基因Br(株高正常)为短节间基因br(植株矮化)的显性。
Br t tbr ttBr T Tbr ttBr T Tbr t t高杆半不育矮杆可育(一)、利用结构变异进行基因定位
2、以易位点为遗传标记(半不育)进行连锁分析(二)、果蝇的ClB测定法(不讲)
倒位杂合体的重组率下降,所以就把倒位染色体上的倒位区段(In)作为抑制交换的显性基因或标志,而把正常染色体作为不能抑制交换的隐性标志。
ClB测定法(Crossover suppress–letha1–Bar technique)正是根据这一点提出的c倒位区段内的一个隐性致死基因可使胚胎在最初发育阶段死亡抑制交换的倒位区段显性棒眼性状,识别倒位X染色体的存在lClB测定法ClB∥ ClB
ClB∥Y
ClB∥X+
雌果蝇死亡雄果蝇死亡雌果蝇活ClB测定就是利用 ClB∥X+雌蝇,测定 X染色体上基因的隐性突变频率 ♀棒眼♂正常眼棒眼正常眼死亡正常眼蓝眼F2♀棒眼♂正常眼棒眼正常眼死亡F1成对杂交棒眼突变性状蓝眼不出现棒眼XClBXClBXClB
XClBAAAAAaAAAaAAAAAaAaaaAXClB
XClB
XClB
X射线XCIB棒眼、显棒眼、显死亡棒眼、显棒眼、显正常眼、显正常眼、显正常眼、显死亡正常眼、显正常眼、显正常眼、隐AaA(二)果蝇的ClB测定(三)、利用易位创造玉米不育系的保持系 ——玉米(四)易位在家蚕生产上的应用(不讲)一、染色体组及其 倍数性
二、同源多倍体
三、异源多倍体
四、多倍体的形成途径及其应用
五、单倍体
六、非整倍体及其应用第2节 染色体数目变异一、染色体组及其倍数性 (一)染色体组(genome)及其基本特征:
染色体基数(x):一个物种染色体组的染色体数目。
染色体组的基本特征:
不同属往往具有独特的染色体基数;
一个染色体组的各个染色体间形态、结构和载有的基因均彼此不同,并且构成一个完整而协调的整体,任何一个成员或其组成部分的缺少对生物都是有害的(生活力降低、配子不育或性状变异)。(二)、整倍体(euploid)整倍体:染色体数目是x的整倍的生物个体
一倍体(monoploid, x) 2n=x
二倍体(diploid, 2x) 2n=2x n=x
三倍体(tripoid, 3x) 2n=3x
四倍体(tetraploid, 4x) 2n=4x n=2x
┆
例:
玉米:二倍体(2n=2x=20, n=x=10)
水稻:二倍体(2n=2x=24, n=x=12)
普通小麦:六倍体(2n=6x=42, n=3x=21, x=7)(二)、整倍体(euploid)多倍体(polyploid):具有三个或三个以上染色体组的整倍体。即:三倍体及以上均称为多倍体。
同源多倍体(autopolyploid) 同源多倍体是指增加的染色体组来自同一物种,一般是由二倍体的染色体直接加倍得到。
异源多倍体(allopolyploid) 异源多倍体是指增加的染色体组来自不同物种,一般是由不同种、属间的杂交种染色体加倍形成的。
二倍体
AA
(2n=2x)二倍体
BB
(2n=2x)同源三倍体
AAA
(2n=3x)同源异源八倍体
AAAABBB
(2n=8x)同源四倍体
AAAA
(2n=4x)异源四倍体
AABB
(2n=4x)染色体加倍染色体加倍染色体加倍(三)、非整倍体(aneuploid)非整倍体:指体细胞核内的染色体不是染色体组的完整倍数,与该物种正常合子(2n)多或少一个以至若干个的现象。
超倍体(hyperploid):染色体数多于2n;
亚倍体(hypoploid):染色体数少于2n。
常见的非整倍体的类型
三体(trisomic): 2n+1
单体(monosomic): 2n-1
双三体(double trisomic): 2n+1+1
双单体(double monosomic): 2n-1-1
四体(tetrasomic): 2n+2
缺体(nullisomic): 2n-2整倍体和非整倍体二、同源多倍体 (一)、同源多倍体的特征形态特征 一般情况下,在一定范围内,随染色体组数增加(也有例外):
◆ 细胞与细胞核体积增大;
◆组织器官(气孔、保卫细胞、叶片、花朵等)巨大化,生物个体更高大粗壮;
◆成熟期延迟、生育期延长。
生理特征
由于基因剂量效应,同源多倍体的生化反应与代谢活动加强;许多性状的表现更强。如:
◆大麦同源四倍体籽粒蛋白质含量比二倍体原种增加10-12%;
◆玉米同源四倍体籽粒胡萝卜素含量比二倍体原种增加43%。
◆大麦白化基因a7不同倍体烟草叶片气孔的比较金鱼草2X金鱼草4X基因剂量 (一)、同源多倍体的特征生殖特征
配子育性降低甚至完全不育。
特殊表型变异
◆基因间平衡与相互作用关系破坏而表现一些异常的性状表现;
◆西葫芦的果形变异:二倍体(梨形)?四倍体(扁圆);
◆菠菜的性别决定:XY型性别决定,四倍体水平只要具有Y染色体(XXXY、XXYY、XYYY、YYYY)就为雄性植株。(二)、同源多倍体的联会与分离1. 同源组与多价体
◆在细胞内,具有同源关系的一组染色体合称为一个同源组
◆二倍体生物细胞内每个同源组有两条同源染色体,减数分裂前期Ⅰ每对同源染色体联会形成一个二价体(Ⅱ)
◆同源多倍体的每个同源组含有三条或三条以上同源染色体,减数分裂前期Ⅰ往往同时有三条以上的染色体参与形成联会复合体,形成多价体(multivalent),如:三价体(Ⅲ)、四价体(Ⅳ)2. 染色体的联会与分离同源多倍体的联会和分离
联会特点: 同源组的同源染色体常联 会成多价体。但是,在任何同源区段 内只能有两条染色体联会,而将其他 染色体的同源区段排斥在联会之外。
因此,每两个染色体之间的只是局部联会,交叉较少,联会松弛,就有可能发生提早解离 2. 染色体的联会与分离——同源三倍体同源三倍体减数分裂和基因分离的特点
同源三倍体在减数分裂前期或形成一个三价体,或形成一个二价体和一个单价体,无论何种配对方式,最后都是一条染色体走向一极,另两条走向一极。
具有两条染色体的配子(2n)的概率是(1/2)n,具有一条染色体的配子(n)的概率也是(1/2)n 。
三倍体所产生的绝大多数配子的染色体数目是在n和2n之间,这些配子的染色体都是不平衡的。不管是哪一种情况,都将造成同源三倍体的配子中染色体组合成分的不平衡,导致同源三倍体的高度不育
农业生产上利用同源三倍体的不育性,生产无籽西瓜(22)、无籽葡萄等(38)
2x
↓
♀ 4x × 2x ♂
↓
3x3. 染色体的联会与分离——同源四倍体玉米4x小孢子: 333个中,n=20为143个,占 42.16%。 其它 n=21~24
n=14~19
玉米4x 与2x 正反交: n多于和小于20也可以参加受精
子代:2n=40 : 60%;
20~39,41~47:40%
育性下降5~10%同源四倍体的联会
同源四倍体联会时可形成一个四价体,或一个三价体和一个单价体,或两个二价体等。
如果同源四倍体的各个染色体在减数分裂中期 I 都以四价体或二价体的构型出现,则后期 I的分离将基本上是均等的,形成的配子多数是可育的。
但多数物种的同源四倍体都是部分不育或高度不育,主要原因是三价体和单价体的频率较高,染色体分配不均衡,造成配子的染色体组成不平衡。同源四倍体:
对于基因(A,a)来讲,二倍体有3种基因型,四倍体有5种基因型。即:AAAA(四式)、AAAa(三式)、AAaa(复式)、Aaaa(单式)、aaaa(零式)。
原理:基因在染色体上距离着丝点的远近决定了同源四倍体的基因分离状况。
3种杂合体分离形成配子的种类和比例,取决于其是按染色体随机分离(基因座位和着丝粒紧密连锁,不发生交换)还是按染色单体随机分离(基因座位和着丝粒距离较远,要考虑交换)。
例如 AAAa基因分离状况?3同源四倍体基因分离—三式(AAAa) 染色体随机分离①.当基因(A-a)染色体上距离着丝点较近,以致基因与着丝点之间很难发生非姊妹染色单体的交换时,则该基因表现染色体随机分离。不同基因配子比例用概率计算 AA配子=C23 =1
Aa配子=C13 × C11=1
AA : Aa =1 : 1
假定这两种精子和卵子都以同样的比率参与受精,则其自交子代的基因型种类和比例为:
(1AA:1Aa)2 = 1AAAA:2AAAa:1AAaa
即全部表现为[A],无[a]
同源四倍体AAaa复式的基因分离同源四倍体染色体随机分离结果3. 同源四倍体染色单体随机分离AAAa②当基因在染色体上距离着丝点较远,基因与着丝点之间发生非姊妹染色单体的交换,导致任何染色单体上的基因可能分配到同一个配子中去,则该基因表现染色单体随机分离, 因此形成配子种类和比例为:
15AA : 12Aa : 1aa
其自交子代:
(15AA:12Aa:1aa)2 = 783[A]:1[a]
同理:
AAaa形成配子为 3AA : 8Aa : 3aa
Aaaa形成配子为 1AA : 12Aa : 15aa同源四倍体染色单体随机分离结果三、异源多倍体异源多倍体是生物进化、新物种形成的重要因素之一
◆被子植物纲中 30-35%
◆禾本科植物 70%
◆许多农作物:小麦、燕麦、甘蔗等
◆其它农作物:烟草、甘蓝型油菜、棉花等
果 树:苹果、梨、草莓
花 卉:菊花、水仙、郁金香
自然界中能正常繁殖的异源多倍体物种几乎都是偶倍数
这类异源多倍体细胞内,每种染色体组都有两个,同源染色体都是成对的,因而减数分裂时能象二倍体一样联会成二价体,所以表现与二倍体相同的性状遗传规律
(一)、偶倍数的异源多倍体 1.偶倍数异源多倍体的形成及证明(人工合成)普通烟草
2n=4x=TTSS=48=24Ⅱ 又称双二倍体
普通小麦
2n=6x=AABBDD=42=21Ⅱ
普通小麦
(T. aestivum)的染色体(一)、偶倍数的异源多倍体2.染色体的部分同源性——部分同源群
◆小麦属染色体的部分同源群(p134)
◆部分同源染色体间可能具有少数相同基因(控制同一性状,表现为多因一效)
◆有时可能相互代替(补偿效应)
◆减数分裂过程中可能发生异源联会(allosynapsis)
A染色体组:1A, 2A, 3A, 4A, 5A, 6A, 7A
B染色体组: lB,2B,3B,4B,5B,6B,7B
D染色体组: lD,2D,3D,4D,5D,6D,7D
编号相同的三组染色体具有部分同源关系,可能有少数基因相同,因而在遗传作用上,有时可以互相替代
同源联会:1A与1A、4B与4B…7D与7D
异源联会:1A与1B或1D…(单倍体ABD)
节段异源多倍体:某异源多倍体的不同染色体组间的部分同源程度很高
有些异源多倍体,由于亲本染色体组的染色体基数不同,自己的各个染色体组的染色体基数也就不同。
例如:
黑芥菜 × 中国油菜
2x=8Ⅱ 2x=10Ⅱ白芥菜 2n=4x=36=8Ⅱ+10Ⅱ=18Ⅱ普通小麦染色体组的部分同源关系染色体组 A B D 黑麦R 大麦H
部分同源组 1A 1B 1D 1R 1H
2A 2B 2D 2R 2H
3A 3B 3D 3R 3H
. . . . .
. . . . .
7A 7B 7D 7R 7H
普通小麦粒色遗传普通小麦粒色
红色对白色为显性
受三对基因控制
R1, r1 3D
R2, r2 3A
R3, r3 3B普通小麦2D与2B的补偿效应 3.染色体组的染色体基数◆偶倍数的异源多倍体是二倍体物种的双二倍体,因此其染色体数是其亲本物种染色体数之和。
◆两亲本物种的染色组的基数可能相同
如:普通烟草(x=12)、普通小麦(x=7)
也可能不同
如芸苔属各物种的染色基数(图)芸苔属(Brassica)各物种的关系(二)、奇倍数的异源多倍体 1 奇倍数异源多倍体的产生及其特征
◆偶倍数异源多倍体物种间杂交
◆奇倍数异源多倍体在联会配对时形成众多的单价体,染色体分离紊乱,配子中染色体组成不平衡,因而很难产生正常可育的配子,所以自然界的物种很难以奇倍数的异源多倍体存在,除非它可以无性繁殖
2.倍半二倍体(sesquidiploid)
形成与用途异源五倍体小麦的形成异源五倍体小麦的联会普通烟草与粘毛烟草的倍半二倍体四、多倍体的形成途径及其应用(一)、未减数配子结合——减数分裂
(二)、体细胞染色体数加倍——有丝分裂
(三)、人工诱导多倍体的应用(一)、未减数配子结合◆桃树(2n=2x=16=8Ⅱ)的未减数配子(n=2x=16)融合形成同源多倍体
未减数配子⊕未减数配子——四倍体(2n=4x=32=8Ⅳ)
未减数配子⊕正常配子 ——三倍体(2n=3x=24=8Ⅲ)
(一)、未减数配子结合◆种间杂种F1未减数配子融合形成异源多倍体
例:(二)、体细胞染色体数加倍体细胞染色体加倍的方法
最常用的方法:秋水仙素处理分生组织
◆阻碍有丝分裂细胞纺锤丝(体)的形成
◆处理浓度:0.01-0.4%(0.2%)
◆处理时间:视材料而定
◆间歇处理效果更好
同源多倍体的诱导
诱导二倍体物种染色体加倍—同源多倍体(偶倍数)
异源多倍体的诱导
◆诱导杂种F1染色体加倍—双二倍体
◆诱导二倍体物种染色体加倍—同源多倍体—杂交—双二倍体(三)、人工诱导多倍体的应用1.克服远缘杂交的不孕性
2.克服远缘杂种的不实性
3.创造种间杂交育种的中间亲本
实质是克服远缘杂交不育性
4.人工合成新物种、育成作物新类型
人工合成同源多倍体
方法:直接加倍
◆例:同源四倍体芥麦、同源三倍体甜菜
人工合成异源多倍体
◆方法:物种间杂交——杂种F1染色体数目加倍
◆实例:八倍体小黑麦(图示) 、六倍体小黑麦(图示)
八倍体小黑麦的人工合成与应用六倍体小黑麦的人工合成与应用五、单倍体(一)、单倍体的类型◆ 单元单倍体(一倍体,n=x):
二倍体生物的单倍体, 就是一倍体.
◆多元单倍体:
四倍体及其以上的偶倍数多倍体所产生的单倍体(具有两个及两个以上的染色体组)。
例如:
六倍体小麦的单倍体具有三个染色体组(n=3x=ABD=21);
四倍体烟草的单倍体具有两个染色体组(n=2x=TS=24)。(二)、单倍体的特点1. 动物: 一般不存在单倍体
例外:雄性单倍体
2.低等生物:生命的主要阶段
3高等植物:高度不育性
◆染色体组成单存在,前期I染色体不能正常联会配对,以单价体形式存在;后期I单价体随机分配或丢失,二分体、四分体染色体组成不完整
◆所有(至少绝大部分)染色单体在后期I进入二分体细胞之一,形成可育配子的机率很小(玉米、小麦)
◆同源四倍体(2n=4x)的单倍体(n=2x)育性水平要高于其它类型玉米单倍体染色体的分离
玉米(2n=2x=20)
其单倍体(n=x=10)主要是以
●5/5、4/6的方式分离
●3/7、2/8、1/9的分离很少
●没有观察到0-10的分离方式。
因而单倍体往往都是高度不育的普通小麦单倍体减数分裂
普通小麦单倍体(n=3x=21=ABD)
减数↓分裂
产生各种染色体组成的配子(0-21)(其中仅具有20,21条染色体的配子具有育性)
配子↓融合
双体(2n=42), 单体(2n=41), 缺体、双单体(2n=40)
双倍体和单倍体的联会特点(三)、单倍体的产生1. 自然产生:由单性生殖产生。
未受精的雌、雄配子,甚至助细胞、反细胞等直接发育形成单倍体胚。
部分动物,如膜翅目昆虫(蜂、蚊)和某些同翅目昆虫(白蚁)的雄性个体都是孤雌生殖形成的自然单倍体。
2. 人工获得单倍体
花药培养:花药离体培养诱导配子体(花粉或子房)发育形成单倍体植株
这是应用最为广泛、成功的人工方法
种间或属间远缘杂交
栽培大麦(Hordeum vudare, 2n=2x=14)与野生球茎大麦(H. bulbosus, 2n=2x=14)杂种胚发育过程中,两物种染色体的行为不协调可导致球茎大麦的染色体逐渐丢失(称为染色体消减现象),可获得大麦的单倍体植株花药培养获得单倍体染色体消减获得单倍体大麦(四)、单倍体在遗传育种研究的应用1.提高育种的选择效率、加速育种进程
基因成单,加倍,纯合体
2.作为良好的遗传研究材料
基因成单,每个基因都发挥作用
3.用以分析染色体组间同源关系
部分同源关系六、非整倍体及其应用非整倍体:染色体数不是染色体组的整倍数的个体
某个体的染色体数比该物种的正常合子染色体数(2n)多或少一个以至若干个染色体。
三体 2n+l=(n–l)Ⅱ+Ⅲ
超倍体 四体 2n+2=(n-1)Ⅱ+Ⅳ
双三体 2n+1+1=(n-2)Ⅱ+2Ⅲ
单体 2n-1=(n-1)Ⅱ+Ⅰ
亚倍体 缺体 2n-2=(n-1)Ⅱ
双单体 2n-1-1=(n-2)Ⅱ+ 2Ⅰ
六、非整倍体及其应用非整倍体的类型
超倍体:多一条或几条染色体,遗传组成不平衡
亚倍体:少一条或几条染色体,遗传物质缺失
非整倍体的形成
减数分裂不正常产生n+1或n-1配子,后代为非整倍体
植物有丝分裂不正常也能产生非整倍体后代
非整倍体的存在
超倍体:二倍体、同源多倍体、异源多倍体均可能
亚倍体:二倍体、同源多倍体、异源多倍体(一)、单体1.单体的特点
●动物:单体的存在往往是许多动物的种性,许多昆虫(蝗虫、蟋蟀)的雌性为 XX型(即2n),雄性为XO型(即2n–l)
●植物:不同植物的单体表现有所不同
二倍体的单体:一般生活力极低而且不育
异源多倍体的单体:具有一定的生活力和育性
普通烟草(2n=4x=TTSS=48)的单体系列
普通小麦(2n=6x=AABBDD=42)的单体系列
(一)、单体普通烟草(2n=4x=TTSS=48)具有24种单体。
●分别用A, B, C, …, V, W, Z字母表示24条染色体;
●24种单体的表示为:
2n-IA, 2n-IB, 2n-IC, …, 2n-IW, 2n-IZ。
●各种单体具有不同的性状变异,表现在:
花冠大小、花萼大小、蒴果大小等性状上。普通小麦的单体系列普通小麦(2n=6x=AABBDD=42)具有21种单体。
●普通小麦的按ABD染色体组及部分同源关系编号为:
A组:1A, 2A, 3A, …, 6A, 7A;
B组:1B, 2B, 3B, …, 6B, 7B;
D组:1D, 2D, 3D, …, 6D, 7D。
●21种单体对应的表示方法为:
2n-I1A, 2n-I2A, …
2n-I1B, 2n-I2B, …
2n-I1D, 2n-I2D, …2.单体染色体的传递普通小麦单体:中期普通小麦单体:终变期硬粒小麦(2n=4x=AABB=28)1A单体2.单体染色体的传递普通小麦单体:四分体普通小麦单体:第一后期普通小麦单体染色体的传递单体自交后代 ●理论上:
◆配子 n: (n-1)=1:1
◆双:单:缺= 1:2:1
●影响因素:
◆单体被遗弃
◆ n, (n-1)参入受精不同
◆ 2n-I, 2n-II 成活率不同
导致不符合理论比例
如
(二)、缺体缺体一般都通过单体自交产生
在异源多倍体生物中可以存在
◆由于缺失一对染色体,对生物个体的性状表现的影响更大,生活力更差
◆普通烟草的缺体在幼胚阶段即死亡
◆普通小麦21种缺体都能够生存
遗传效应
◆表现广泛的性状变异(例)
◆通过缺体的性状变异,可能确定位于该染色体上的基因普通小麦缺体系列的穗形(三)、三体1.三体的性状变异
不同物种,不同染色体的三体的变异性状及程度不同
◆直果曼陀罗(2n=2x=24)的果型变异(图)
◆玉米(2n=2x=20)有10个不同的三体
◆普通小麦(2n=2x=42)具有21个三体,但性状变异较小
◆人类: XXX, XXY XYY三体的染色体联会与分离◆三体染色体联会与分离(图1、2、3、4、5、6)
(1)Ⅲ 2/1
(2)Ⅱ+Ⅰ 2/1 (单价体随机进入一个二分体细胞)
1/1 (单价体丢失)
◆形成四分体细胞:
(1)种类:n, n+1
(2)比例:n>n+1
◆配子育性与受精结合
配子育性:n>n+1, 尤其是在花粉中,因此n+1配子主要通过雌配子传递
后代: 小麦 双体(54.1%),三体(45%),四体(1%)
玉米 ♂ 三体1.4% 双体98.6%
♀ 三体24.5% 双体75.5%三体终变期:链式三价体三体染色体联会三体后期 I:2/1式分离三体末期 I:落后三价体复式三体的基因分离a 染色体随机分离(AAa)
(n+1) n
1,2/3 → AA AA a a
AAa → 1,3/2 → Aa Aa A A
2,3/1 → Aa Aa A A
AA:Aa:A:a=1:2:2:1
若n+l配子与n配子同等可育,且精子和卵子也同等可育,则自交子代的表现型比例就应该是[A]:[a]=35:1复式三体的基因分离假定复式三体的n+l精子全部不能参与受精,参与受精的卵子中1/4是n+l,3/4是n,则自交:
♀ ♂ 2A 1a
1AA
2Aa [A]:[a]=11:1
6A
3a复式三体染色体随机分离复式三体染色单体随机分离 ◆配子
n配子: A:4;a:2 (10:5)
n+1配子:AA:C42 = 6;
Aa: C41 C21 = 8;
aa: C21 = 1 (14:1)
配子:A:a = 24:1
◆表型
母本:3/4 (n+1)配子受精;
父本 (n+1)配子不授精;
表型:[A]:[a] = 10.3:1
b 染色单体随机分离(AAa)
分离的单位为6个染色单体:4A,2a
三价体的分离主要是2/1式,形成(n+1)和n
4!
则 AA配子=C24= = 6
(4-2)!2!
4! 2!
Aa配子=C14?C12= ? = 8
(4-1)!1! (2-1)!1!
2!
aa配子=C22= = 1
(2-2)!2!
即 AA : Aa : aa = 6:8:1
n配子与n+1配子数相等,即1(n+1):1n
n配子中 A:a=C14:C12=2:1=10:5
则全部配子种类和比例:
6AA:8Aa:1aa:10A:5a
若n+1配子与n配子同等可育,♀♂同等
参与受精,则自交子代:
(6AA:8Aa:1aa:10A:5a)2 = 24[A]:1[a]若参与受精的♀中3/4是n,1/4是n+1
n+1精子全部不能参与受精
则自交子代:
(6AA:8Aa:1aa:30A:15a)(2A:1a)
= 10.25[A]:1[a]
若同时考虑(n-1)Ⅱ中某一对基因如Bb
则自交子代:
(3B:1b)(10.25A:1a)(四)、四体与同源四倍体相比
◆只有一个同源组具有四条染色体
◆后期 I 2/2式分离的比例更高
四体小麦自交:四体73.8%, 三体23.6%, 双体1.9%
◆基因的分离与同源四倍体类似
◆生活力和配子的育性均更高(五)、非整倍体的应用1.基因的染色体定位(1).利用单体进行基因定位
隐性基因定位
普通烟草黄绿突变(yg2)的定位(过程)
单体定位机理
a表现双体与对应单体杂交(图)
a表现双体与非对应单体杂交(图)
显性基因定位
基本过程(p144)普通烟草黄绿突变(yg2)的定位普通烟草的黄绿突变型基因(隐性yg2)定位:
绿叶单体(2n-Ix)(共24种)×黄绿色叶型双体(yg2yg2)
↓
杂种F1(共24种)
(考察F1性状表现)
23种F1均表现为绿叶 仅与[2n-Is]杂种F1表现为:
绿叶和黄绿叶两种
(检查F1各个体的染色体数目)
所有绿叶个体均为双体(2n) yg2基因位于S染色体上?黄绿叶个体均为单体(2n-1)当隐性基因(a)在某染色体上时,则
P ((n–1)Ⅱ+ⅠA) × ((n–1)Ⅱ+Ⅱaa)
↙ ↘ ↓
n n–1 n
G (n–1) I+IA (n–1)I (n–1)I+Ia
↓ ↓
2n 2n–1
F1 ((n–1)Ⅱ+ⅡAa) ((n–1) Ⅱ+Ia)
A表现型 a表现型
双体 单体当隐性基因(a)不在某染色体上时,则
P ((n–1)ⅡAA +Ⅰ) × ((n–1)Ⅱaa +Ⅱ)
↙ ↘ ↓
n n–1 n
G (n–1) IA+I (n–1)IA (n–1)Ia+I
↓ ↓
2n 2n–1
F1 ((n–1)ⅡAa +Ⅱ) ((n–1) ⅡAa +I) 双体单体A表现型 A表现型a表现双体与对应单体杂交a表现双体与非对应单体杂交②单体测验显性基因
若显性基因(A)在某染色体上时,则F1单体自交后代中只有缺体是a表型
P ((n–1)Ⅱ+Ⅰa) × ((n–1)Ⅱ +ⅡAA )
↓
F1 ((n–1)Ⅱ+ⅡAa) ((n–1)Ⅱ+IA)
↓
♀ ♂ (n-1)Ⅰ+ IA (n-1) Ⅰ
F2 (n–1)Ⅱ+ⅡAA(n–1)Ⅱ+IA(n–1)Ⅱ+IA(n–1)Ⅱ 表现[a](n-1)Ⅰ+ IA(n-1) Ⅰ?当显性基因(A)不在某染色体上时,则其F1单体所自交的F2群体内,双体、单体、缺体植株都会有少数是a表现型的。
显性基因的单体定位过程隐性单体系列(n种)×显性纯合双体(AA)
↓
杂种F1(均表现为显性)(n种)
(进行染色体数目鉴定)
(单体↓自交)
F2(n种)
(鉴定性状表现、鉴定隐性F2的染色体数目)
n-1种隐性F2含双体、单体、缺体各种类型
只有1种F2双体、单体均为显性?基因所在1.基因的染色体定位(2).利用三体进行隐性基因定位
双体
自交后代表现型比例=3显:1隐
测交后代表现型比例=1显:1隐
三体
自交后代表现型比例≠3显:1隐
测交后代表现型比例≠1显:1隐② 三体测验
AAA×aa
↓
F1 AAa
↓
F2 所要测验的基因不在三体染色体上,
那么F2应该出现3A:1a
所要测验的基因在三体染色体上,
那么F2应该出现35A:1a或其他比例 ?(2)有目标地替换染色体
用单体有目标地替换染色体
已知某抗病基因R在小麦的6B染色体上,某优良品种不抗病(6BIIrr),要替换成抗病品种的6BIIRR:
(20II+6BIr) × (20II+6BIIRR)
?
F1 (20II+6BIIRr) (20II+6BIR)
?
F2 (20II+6BIIRR) 用单体并通过倍半二倍体有目标地替换染色体
使小麦只换取黑麦的个别染色体:
AABBDDRR × AABBDD
?
F1 AABBDDR
?
F2 (AABBDD+IR) × (AABBDD-IT)
?
20II+IR+IT
?
2n=20IITT+IIRR=21II用缺体有目标地替换染色体
已知某抗病基因R在小麦的6D染色体上,某优良品种不抗病(6DIIrr),要替换成抗病品种的6BIIRR:
(21II-6DII) × (20II+6DIIRR)
?
F1 (20II + 6DIR)
?
F2 (20II + 6DIIRR) 2.有目标地替换染色体小麦抗病品种: 20II+6BIIRR;
不抗病6B单体: 20II+ 6BIIrr;
20II+ 6BIr;x 20II+6BIIRR
20II+6BIIRr(双体抗病) 20II+6BIR (单体抗病)
?
20II+6BIIRR
小麦:
AABBDDRR x AABBDD
AABBDD+R(7I)
(2n+1): AABBDD+I1R
AABBDD+I2R
…….…
ABD+IR x ABD-IT
20II+IT+IR
?
(异代换系) 20IIT+IIR
异附加系:
AABBDD+II1R
……………
缺体
21II – 6DII x 20II+6DIIrr
20II+6DIR
?
20II+6DIIRR
?
第2节 染色体数目变异
第一节 染色体结构变异结构变异的形成:断裂—重接
◎使染色体产生折断的因素:
自然:温度剧变、营养生理条件异常、遗传因素等;
人为:物理射线与化学药剂处理等。
◎染色体折断的结果:
正确重接:重新愈合,恢复原状;
错误重接:产生结构变异;
保持断头:产生结构变异。
◎结构变异的基本类型:
缺失、重复、倒位、易位
概念、类型与特点;
细胞学特征与鉴定;
遗传效应。第一节 染色体结构变异◎ 缺失
◎ 重复
◎ 倒位
◎ 易位
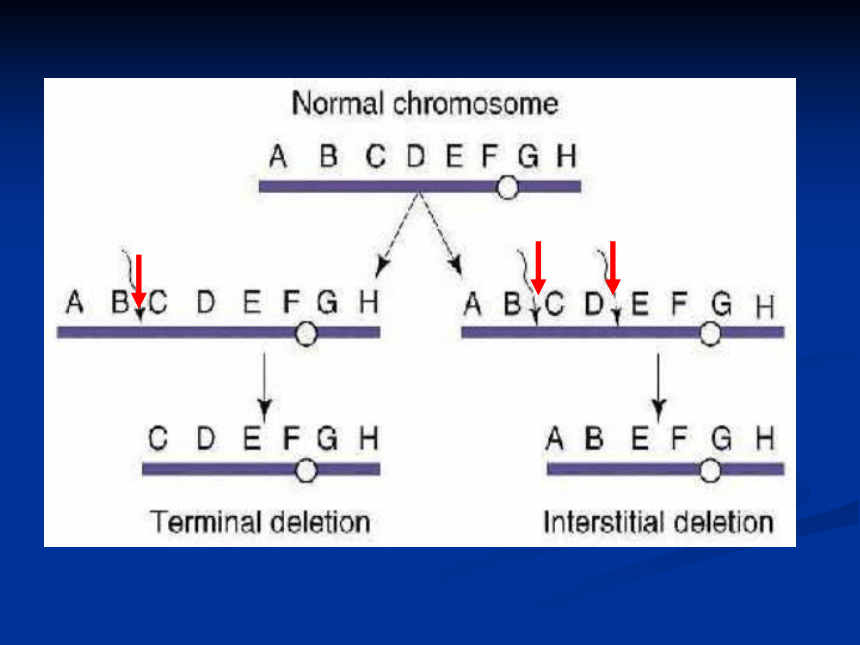
◎ 染色体结构变异的应用(一)、缺失 的类型
缺失:染色体的某一区段丢失了
断片:缺失的区段无着丝粒
顶端缺失:缺失的区段为某臂的外端
某一整臂缺失了就成为顶端着丝点染色体
中间缺失:缺失的区段为某臂的内段 顶端缺失染色体很难定型,常会重接。
(1)断头很难愈合,断头可能同另一有着丝粒的染色体的断头重接,成为单着粒染色体
(2)顶端缺失染色体的两个姊妹染色单体可能在断头上彼此接合,形成双着丝粒染色体
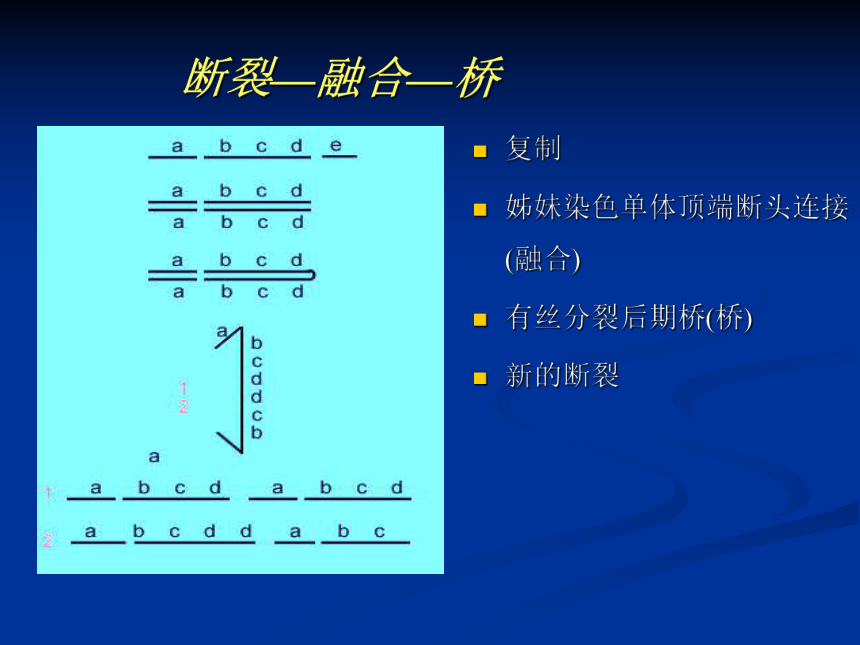
双着丝粒染色体就会在细胞分裂的后期受两个着丝粒向相反两极移动所产生的拉力所折断,再次造成结构的变异而不能稳定,形成断裂—融合—桥 断裂—融合—桥复制
姊妹染色单体顶端断头连接(融合)
有丝分裂后期桥(桥)
新的断裂
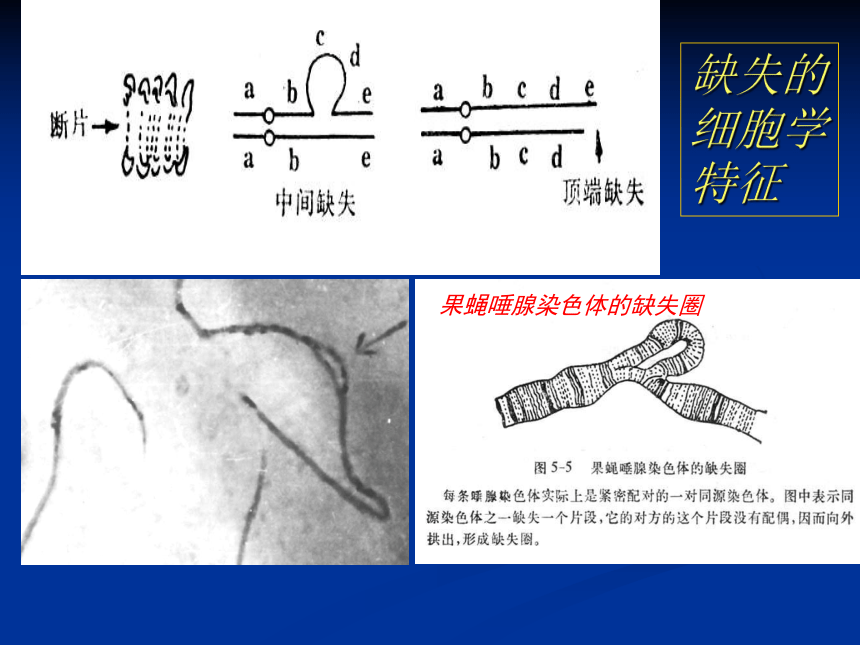
(二)、缺失的细胞学鉴定无着丝粒断片;
最初发生缺失的细胞在分裂时可见无着丝粒断片。
缺失环(环形或瘤形突出);
中间缺失杂合体偶线期和粗线期出现;
二价体末端突出;
顶端缺失杂合体粗线期、双线期,交叉未完全端化的二价体末端不等长。缺失的细胞学特征果蝇唾腺染色体的缺失圈(三)、缺失的遗传效应缺失纯合体:
致死或半致死。
缺失杂合体:
缺失区段较长时,生活力差、配子(尤其是花粉)败育或竞争不过正常配子;
缺失区段较小时,可能会造成假显性现象或其它异常现象(p117例:猫叫综合症)。缺失区段上基因丢失导致:
基因所决定、控制的生物功能丧失或异常;
基因间相互作用关系破坏;
基因排列位置关系改变。
影响缺失对生物个体危害程度的因素:
缺失区段的大小;
缺失区段所含基因的多少;
缺失基因的重要程度;
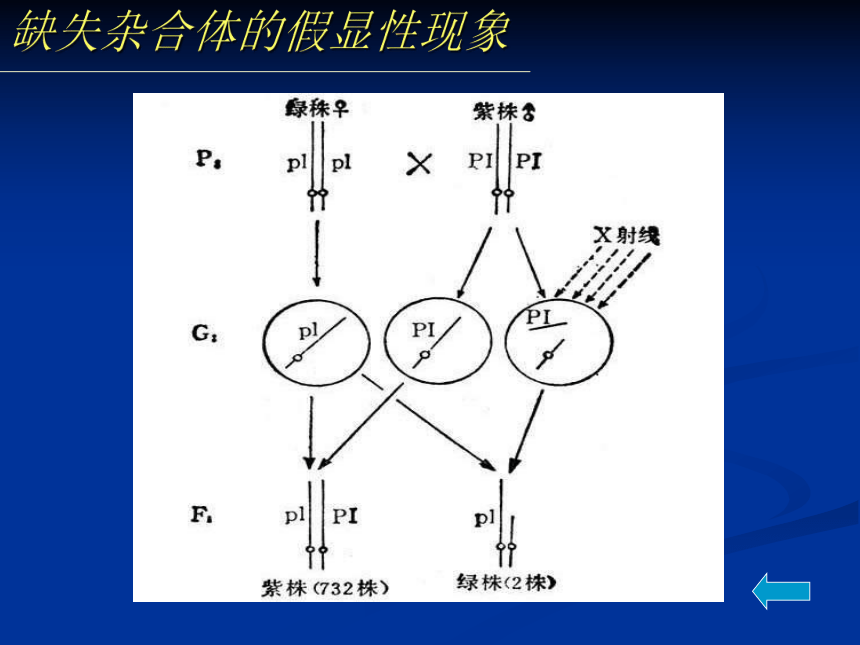
染色体倍性水平。缺失杂合体的假显性现象猫叫综合症 第5号染色体缺失(短臂缺失)患儿发
出咪咪声,耳位低下,智商仅20~40.猫叫综合症“猫叫综合症”是第5号染色体短臂缺失引起的遗传病,发生率为十万分之一,在国内外均很少见。患儿一般表现为生长发育迟缓,头央部畸形,哭声轻,音调高,皮纹改变等特点,并有严重的智能障碍,而其最明显的特征是哭声类似猫叫,“猫叫综合症”因此而得名。据称,病儿哭声异常可能系喉部发育不良所致,也可能与脑损害有关。 二、重复(duplication)(一)、重复的类别与形成
重复:染色体多了自己的某一区段。
顺接重复(tanden duplication) ;
反接重复(reverse duplication) 。(二)、重复的细胞学鉴定重复圈(环)果蝇X染色体上16A区段重复的形成 重复区段内不能有着丝粒,否则重复染色体就变成双着丝粒的染色体,就会继续发生结构变异,很难稳定成型。
重复和缺失总是伴随出现的。某染色体的一个区段转移给同源的另一个染色体之后,它自己就成为缺失染色体了。 (三)、重复的遗传效应重复对个体综合表现的影响:
重复区段内的基因重复,影响基因间的平衡关系;
会影响个体的生活力(影响的程度与重复区段的大小有关)。
剂量效应(dosage effect):
果蝇眼色遗传的剂量效应,红色(V+)对朱红(V)为显性;
杂合体(V+V)表现为红色;
但(V+VV)的表现型却为朱红色。
位置效应(position effect):
果蝇眼面大小遗传的位置效应;
位置效应的意义。果蝇棒眼的位置效应三、倒位(inversion)(一)、倒位的类别与形成
倒位:染色体某区段的正常直线顺序颠倒了
臂内倒位(paracentric inversion);
臂间倒位(pericentic inversion)。(二)、倒位的细胞学鉴定倒位杂合体的联会倒位杂合体的“倒位圈”(三)、倒位的遗传效应1. 倒位杂合体的部分不育现象:
倒位圈内发生交换后,产生的交换型配子(50%)含重复缺失染色单体,这类配子是不育的;
只有部分孢母细胞在减数分裂时倒位圈内会发生非姊妹染色单体间的交换;
倒位点可以当作一个显性基因位点看待,其性状表现就是倒位杂合体部分不育。
2. 倒位改变了基因在染色体上的排列:
基因间距离关系发生改变;
可能引起倒位区段基因的位置效应;
倒位杂合体的基因间交换值降低:
倒位圈的结构影响联会复合体的正常形成;
倒位圈内发生交换后产生的交换型配子是不育的。
3. 是物种进化的重要因素之一。
倒位可能导致新物种的产生。3、倒位的遗传效应 (1)位置效应:倒位区段内、外各个基因之间的物理距离发生改变,其遗传距离一般也改变。 5a倒位杂合体的重组率(Pr-bt1)0.4 %、(bm1-bt1)0%、(a2-bt1)4.5%(2)降低倒位杂合体上连锁基因的重组率:产生的交换型配子数明显减少,故重组率降低。 (3)倒位可以形成新种,促进生物进化例如:果蝇(n=4):不同倒位特点的种,分布在不同地理区域。倒位杂合体的交换臂间倒位杂合体的交换臂内倒位杂合体的交换臂内倒位形成的“后期 I 桥”四、易位(translocation)(一)、易位的类别与形成
相互易位(reciprocal translocation)
简单易位(simple translocation)(转移)
易位的形成(
易位染色体
易位染色体的表示方法
易位杂合体(translocation heterozygote)
易位纯合体(translocation homozygote)1221112212易位的类别与形成(二)、易位的细胞学鉴定易位杂合体的联会和分离易位杂合体的联会和分离1,121221121,122,212,21全部不育配子相邻式分离11122212,122,121,211,21全部不育配子相邻式分离2全部可育配子交替式分离易位杂合体粗线期十字形配对易位杂合体的联会和分离玉米(2n=20)终变期染色体(三)、易位的遗传效应易位杂合体半不育现象
易位杂合体十字形配对进行交替式分离所产生的配子是可育的,而进行相邻式分离产生的配子是不育的。
花粉50%败育,胚囊50%败育,结实率只有50%,由半不育植株的种子所长出的植株又会有半数是半不育的,半数是正常可育的。
易位杂合体后期分离时:
1,21/12,2 相邻式,重复缺失-败育,1/2
1,12/2,21
1,2/12,21 交替式,正常/易位-可育,1/2(三)、易位的遗传效应易位杂合体半不育现象
玉米型:相邻式与交换式各占50%,配子半不育,可视为一个半不育显性基因看待。
月见草型:全部采用交替式分离,所有配子都可育。
曼驼罗、风铃草、紫万年青, 易位杂合体后期分离100%是交替式 。
(三)、易位的遗传效应易位改变了生物的连锁群
基因间连锁关系和位置效应变化
易位杂合体的基因重组值降低
十字形结构影响联会复合体中交换的正常形成
易位与生物进化和新物种形成
相互易位纯合体可以保存形成的新的连锁群关系
易位可能导致染色体融合,引起染色体数目变异染色体融合易位造成染色体融合,导致染色体数目的变异
AB丢失Robertsonian translocation五、 染色体结构变异的应用(一)、利用结构变异进行基因定位
利用缺失的假显性现象将基因定位到染色体某一个区段
以易位点为遗传标记(半不育)进行连锁分析
(二)、果蝇的ClB测定
(三)、利用易位创造玉米不育系的保持系
(四)、易位在家蚕生产上的应用1、利用缺失造成的假显性现象,可以进行基因定位
使载有显性基因的染色体发生缺失,让其隐性等位基因表现“假显性” 对表现假显性的个体进行细胞学鉴定,发现某染色体缺失了某一区段,就说明该显性基因位于该染色体的缺失区段上。 (一)、利用结构变异进行基因定位(一)、利用结构变异进行基因定位
2、以易位点为遗传标记(半不育)进行连锁分析易位杂交体产生的可育配子中一半含两个正常染色体(1,2),一半含两个易位染色体(12,21)。那么在易位杂合体自交子代群体内, 1/4正常可育个体, 2/4半不育易位杂合体, 1/4可育易位纯合体
易位染色体的易位接合点相当于一个半不育的显性遗传基因(T),正常染色体与易位接合点相对的等位点,则相当于一个可育的隐性遗传基因(t)
遗传学上利用这一特点,用两点或三点测验,根据T–t与某邻近基因之间的重组率,确定易位接合点在染色体上的位置 。
例:玉米长节间基因Br(株高正常)为短节间基因br(植株矮化)的显性。
Br t tbr ttBr T Tbr ttBr T Tbr t t高杆半不育矮杆可育(一)、利用结构变异进行基因定位
2、以易位点为遗传标记(半不育)进行连锁分析(二)、果蝇的ClB测定法(不讲)
倒位杂合体的重组率下降,所以就把倒位染色体上的倒位区段(In)作为抑制交换的显性基因或标志,而把正常染色体作为不能抑制交换的隐性标志。
ClB测定法(Crossover suppress–letha1–Bar technique)正是根据这一点提出的c倒位区段内的一个隐性致死基因可使胚胎在最初发育阶段死亡抑制交换的倒位区段显性棒眼性状,识别倒位X染色体的存在lClB测定法ClB∥ ClB
ClB∥Y
ClB∥X+
雌果蝇死亡雄果蝇死亡雌果蝇活ClB测定就是利用 ClB∥X+雌蝇,测定 X染色体上基因的隐性突变频率 ♀棒眼♂正常眼棒眼正常眼死亡正常眼蓝眼F2♀棒眼♂正常眼棒眼正常眼死亡F1成对杂交棒眼突变性状蓝眼不出现棒眼XClBXClBXClB
XClBAAAAAaAAAaAAAAAaAaaaAXClB
XClB
XClB
X射线XCIB棒眼、显棒眼、显死亡棒眼、显棒眼、显正常眼、显正常眼、显正常眼、显死亡正常眼、显正常眼、显正常眼、隐AaA(二)果蝇的ClB测定(三)、利用易位创造玉米不育系的保持系 ——玉米(四)易位在家蚕生产上的应用(不讲)一、染色体组及其 倍数性
二、同源多倍体
三、异源多倍体
四、多倍体的形成途径及其应用
五、单倍体
六、非整倍体及其应用第2节 染色体数目变异一、染色体组及其倍数性 (一)染色体组(genome)及其基本特征:
染色体基数(x):一个物种染色体组的染色体数目。
染色体组的基本特征:
不同属往往具有独特的染色体基数;
一个染色体组的各个染色体间形态、结构和载有的基因均彼此不同,并且构成一个完整而协调的整体,任何一个成员或其组成部分的缺少对生物都是有害的(生活力降低、配子不育或性状变异)。(二)、整倍体(euploid)整倍体:染色体数目是x的整倍的生物个体
一倍体(monoploid, x) 2n=x
二倍体(diploid, 2x) 2n=2x n=x
三倍体(tripoid, 3x) 2n=3x
四倍体(tetraploid, 4x) 2n=4x n=2x
┆
例:
玉米:二倍体(2n=2x=20, n=x=10)
水稻:二倍体(2n=2x=24, n=x=12)
普通小麦:六倍体(2n=6x=42, n=3x=21, x=7)(二)、整倍体(euploid)多倍体(polyploid):具有三个或三个以上染色体组的整倍体。即:三倍体及以上均称为多倍体。
同源多倍体(autopolyploid) 同源多倍体是指增加的染色体组来自同一物种,一般是由二倍体的染色体直接加倍得到。
异源多倍体(allopolyploid) 异源多倍体是指增加的染色体组来自不同物种,一般是由不同种、属间的杂交种染色体加倍形成的。
二倍体
AA
(2n=2x)二倍体
BB
(2n=2x)同源三倍体
AAA
(2n=3x)同源异源八倍体
AAAABBB
(2n=8x)同源四倍体
AAAA
(2n=4x)异源四倍体
AABB
(2n=4x)染色体加倍染色体加倍染色体加倍(三)、非整倍体(aneuploid)非整倍体:指体细胞核内的染色体不是染色体组的完整倍数,与该物种正常合子(2n)多或少一个以至若干个的现象。
超倍体(hyperploid):染色体数多于2n;
亚倍体(hypoploid):染色体数少于2n。
常见的非整倍体的类型
三体(trisomic): 2n+1
单体(monosomic): 2n-1
双三体(double trisomic): 2n+1+1
双单体(double monosomic): 2n-1-1
四体(tetrasomic): 2n+2
缺体(nullisomic): 2n-2整倍体和非整倍体二、同源多倍体 (一)、同源多倍体的特征形态特征 一般情况下,在一定范围内,随染色体组数增加(也有例外):
◆ 细胞与细胞核体积增大;
◆组织器官(气孔、保卫细胞、叶片、花朵等)巨大化,生物个体更高大粗壮;
◆成熟期延迟、生育期延长。
生理特征
由于基因剂量效应,同源多倍体的生化反应与代谢活动加强;许多性状的表现更强。如:
◆大麦同源四倍体籽粒蛋白质含量比二倍体原种增加10-12%;
◆玉米同源四倍体籽粒胡萝卜素含量比二倍体原种增加43%。
◆大麦白化基因a7不同倍体烟草叶片气孔的比较金鱼草2X金鱼草4X基因剂量 (一)、同源多倍体的特征生殖特征
配子育性降低甚至完全不育。
特殊表型变异
◆基因间平衡与相互作用关系破坏而表现一些异常的性状表现;
◆西葫芦的果形变异:二倍体(梨形)?四倍体(扁圆);
◆菠菜的性别决定:XY型性别决定,四倍体水平只要具有Y染色体(XXXY、XXYY、XYYY、YYYY)就为雄性植株。(二)、同源多倍体的联会与分离1. 同源组与多价体
◆在细胞内,具有同源关系的一组染色体合称为一个同源组
◆二倍体生物细胞内每个同源组有两条同源染色体,减数分裂前期Ⅰ每对同源染色体联会形成一个二价体(Ⅱ)
◆同源多倍体的每个同源组含有三条或三条以上同源染色体,减数分裂前期Ⅰ往往同时有三条以上的染色体参与形成联会复合体,形成多价体(multivalent),如:三价体(Ⅲ)、四价体(Ⅳ)2. 染色体的联会与分离同源多倍体的联会和分离
联会特点: 同源组的同源染色体常联 会成多价体。但是,在任何同源区段 内只能有两条染色体联会,而将其他 染色体的同源区段排斥在联会之外。
因此,每两个染色体之间的只是局部联会,交叉较少,联会松弛,就有可能发生提早解离 2. 染色体的联会与分离——同源三倍体同源三倍体减数分裂和基因分离的特点
同源三倍体在减数分裂前期或形成一个三价体,或形成一个二价体和一个单价体,无论何种配对方式,最后都是一条染色体走向一极,另两条走向一极。
具有两条染色体的配子(2n)的概率是(1/2)n,具有一条染色体的配子(n)的概率也是(1/2)n 。
三倍体所产生的绝大多数配子的染色体数目是在n和2n之间,这些配子的染色体都是不平衡的。不管是哪一种情况,都将造成同源三倍体的配子中染色体组合成分的不平衡,导致同源三倍体的高度不育
农业生产上利用同源三倍体的不育性,生产无籽西瓜(22)、无籽葡萄等(38)
2x
↓
♀ 4x × 2x ♂
↓
3x3. 染色体的联会与分离——同源四倍体玉米4x小孢子: 333个中,n=20为143个,占 42.16%。 其它 n=21~24
n=14~19
玉米4x 与2x 正反交: n多于和小于20也可以参加受精
子代:2n=40 : 60%;
20~39,41~47:40%
育性下降5~10%同源四倍体的联会
同源四倍体联会时可形成一个四价体,或一个三价体和一个单价体,或两个二价体等。
如果同源四倍体的各个染色体在减数分裂中期 I 都以四价体或二价体的构型出现,则后期 I的分离将基本上是均等的,形成的配子多数是可育的。
但多数物种的同源四倍体都是部分不育或高度不育,主要原因是三价体和单价体的频率较高,染色体分配不均衡,造成配子的染色体组成不平衡。同源四倍体:
对于基因(A,a)来讲,二倍体有3种基因型,四倍体有5种基因型。即:AAAA(四式)、AAAa(三式)、AAaa(复式)、Aaaa(单式)、aaaa(零式)。
原理:基因在染色体上距离着丝点的远近决定了同源四倍体的基因分离状况。
3种杂合体分离形成配子的种类和比例,取决于其是按染色体随机分离(基因座位和着丝粒紧密连锁,不发生交换)还是按染色单体随机分离(基因座位和着丝粒距离较远,要考虑交换)。
例如 AAAa基因分离状况?3同源四倍体基因分离—三式(AAAa) 染色体随机分离①.当基因(A-a)染色体上距离着丝点较近,以致基因与着丝点之间很难发生非姊妹染色单体的交换时,则该基因表现染色体随机分离。不同基因配子比例用概率计算 AA配子=C23 =1
Aa配子=C13 × C11=1
AA : Aa =1 : 1
假定这两种精子和卵子都以同样的比率参与受精,则其自交子代的基因型种类和比例为:
(1AA:1Aa)2 = 1AAAA:2AAAa:1AAaa
即全部表现为[A],无[a]
同源四倍体AAaa复式的基因分离同源四倍体染色体随机分离结果3. 同源四倍体染色单体随机分离AAAa②当基因在染色体上距离着丝点较远,基因与着丝点之间发生非姊妹染色单体的交换,导致任何染色单体上的基因可能分配到同一个配子中去,则该基因表现染色单体随机分离, 因此形成配子种类和比例为:
15AA : 12Aa : 1aa
其自交子代:
(15AA:12Aa:1aa)2 = 783[A]:1[a]
同理:
AAaa形成配子为 3AA : 8Aa : 3aa
Aaaa形成配子为 1AA : 12Aa : 15aa同源四倍体染色单体随机分离结果三、异源多倍体异源多倍体是生物进化、新物种形成的重要因素之一
◆被子植物纲中 30-35%
◆禾本科植物 70%
◆许多农作物:小麦、燕麦、甘蔗等
◆其它农作物:烟草、甘蓝型油菜、棉花等
果 树:苹果、梨、草莓
花 卉:菊花、水仙、郁金香
自然界中能正常繁殖的异源多倍体物种几乎都是偶倍数
这类异源多倍体细胞内,每种染色体组都有两个,同源染色体都是成对的,因而减数分裂时能象二倍体一样联会成二价体,所以表现与二倍体相同的性状遗传规律
(一)、偶倍数的异源多倍体 1.偶倍数异源多倍体的形成及证明(人工合成)普通烟草
2n=4x=TTSS=48=24Ⅱ 又称双二倍体
普通小麦
2n=6x=AABBDD=42=21Ⅱ
普通小麦
(T. aestivum)的染色体(一)、偶倍数的异源多倍体2.染色体的部分同源性——部分同源群
◆小麦属染色体的部分同源群(p134)
◆部分同源染色体间可能具有少数相同基因(控制同一性状,表现为多因一效)
◆有时可能相互代替(补偿效应)
◆减数分裂过程中可能发生异源联会(allosynapsis)
A染色体组:1A, 2A, 3A, 4A, 5A, 6A, 7A
B染色体组: lB,2B,3B,4B,5B,6B,7B
D染色体组: lD,2D,3D,4D,5D,6D,7D
编号相同的三组染色体具有部分同源关系,可能有少数基因相同,因而在遗传作用上,有时可以互相替代
同源联会:1A与1A、4B与4B…7D与7D
异源联会:1A与1B或1D…(单倍体ABD)
节段异源多倍体:某异源多倍体的不同染色体组间的部分同源程度很高
有些异源多倍体,由于亲本染色体组的染色体基数不同,自己的各个染色体组的染色体基数也就不同。
例如:
黑芥菜 × 中国油菜
2x=8Ⅱ 2x=10Ⅱ白芥菜 2n=4x=36=8Ⅱ+10Ⅱ=18Ⅱ普通小麦染色体组的部分同源关系染色体组 A B D 黑麦R 大麦H
部分同源组 1A 1B 1D 1R 1H
2A 2B 2D 2R 2H
3A 3B 3D 3R 3H
. . . . .
. . . . .
7A 7B 7D 7R 7H
普通小麦粒色遗传普通小麦粒色
红色对白色为显性
受三对基因控制
R1, r1 3D
R2, r2 3A
R3, r3 3B普通小麦2D与2B的补偿效应 3.染色体组的染色体基数◆偶倍数的异源多倍体是二倍体物种的双二倍体,因此其染色体数是其亲本物种染色体数之和。
◆两亲本物种的染色组的基数可能相同
如:普通烟草(x=12)、普通小麦(x=7)
也可能不同
如芸苔属各物种的染色基数(图)芸苔属(Brassica)各物种的关系(二)、奇倍数的异源多倍体 1 奇倍数异源多倍体的产生及其特征
◆偶倍数异源多倍体物种间杂交
◆奇倍数异源多倍体在联会配对时形成众多的单价体,染色体分离紊乱,配子中染色体组成不平衡,因而很难产生正常可育的配子,所以自然界的物种很难以奇倍数的异源多倍体存在,除非它可以无性繁殖
2.倍半二倍体(sesquidiploid)
形成与用途异源五倍体小麦的形成异源五倍体小麦的联会普通烟草与粘毛烟草的倍半二倍体四、多倍体的形成途径及其应用(一)、未减数配子结合——减数分裂
(二)、体细胞染色体数加倍——有丝分裂
(三)、人工诱导多倍体的应用(一)、未减数配子结合◆桃树(2n=2x=16=8Ⅱ)的未减数配子(n=2x=16)融合形成同源多倍体
未减数配子⊕未减数配子——四倍体(2n=4x=32=8Ⅳ)
未减数配子⊕正常配子 ——三倍体(2n=3x=24=8Ⅲ)
(一)、未减数配子结合◆种间杂种F1未减数配子融合形成异源多倍体
例:(二)、体细胞染色体数加倍体细胞染色体加倍的方法
最常用的方法:秋水仙素处理分生组织
◆阻碍有丝分裂细胞纺锤丝(体)的形成
◆处理浓度:0.01-0.4%(0.2%)
◆处理时间:视材料而定
◆间歇处理效果更好
同源多倍体的诱导
诱导二倍体物种染色体加倍—同源多倍体(偶倍数)
异源多倍体的诱导
◆诱导杂种F1染色体加倍—双二倍体
◆诱导二倍体物种染色体加倍—同源多倍体—杂交—双二倍体(三)、人工诱导多倍体的应用1.克服远缘杂交的不孕性
2.克服远缘杂种的不实性
3.创造种间杂交育种的中间亲本
实质是克服远缘杂交不育性
4.人工合成新物种、育成作物新类型
人工合成同源多倍体
方法:直接加倍
◆例:同源四倍体芥麦、同源三倍体甜菜
人工合成异源多倍体
◆方法:物种间杂交——杂种F1染色体数目加倍
◆实例:八倍体小黑麦(图示) 、六倍体小黑麦(图示)
八倍体小黑麦的人工合成与应用六倍体小黑麦的人工合成与应用五、单倍体(一)、单倍体的类型◆ 单元单倍体(一倍体,n=x):
二倍体生物的单倍体, 就是一倍体.
◆多元单倍体:
四倍体及其以上的偶倍数多倍体所产生的单倍体(具有两个及两个以上的染色体组)。
例如:
六倍体小麦的单倍体具有三个染色体组(n=3x=ABD=21);
四倍体烟草的单倍体具有两个染色体组(n=2x=TS=24)。(二)、单倍体的特点1. 动物: 一般不存在单倍体
例外:雄性单倍体
2.低等生物:生命的主要阶段
3高等植物:高度不育性
◆染色体组成单存在,前期I染色体不能正常联会配对,以单价体形式存在;后期I单价体随机分配或丢失,二分体、四分体染色体组成不完整
◆所有(至少绝大部分)染色单体在后期I进入二分体细胞之一,形成可育配子的机率很小(玉米、小麦)
◆同源四倍体(2n=4x)的单倍体(n=2x)育性水平要高于其它类型玉米单倍体染色体的分离
玉米(2n=2x=20)
其单倍体(n=x=10)主要是以
●5/5、4/6的方式分离
●3/7、2/8、1/9的分离很少
●没有观察到0-10的分离方式。
因而单倍体往往都是高度不育的普通小麦单倍体减数分裂
普通小麦单倍体(n=3x=21=ABD)
减数↓分裂
产生各种染色体组成的配子(0-21)(其中仅具有20,21条染色体的配子具有育性)
配子↓融合
双体(2n=42), 单体(2n=41), 缺体、双单体(2n=40)
双倍体和单倍体的联会特点(三)、单倍体的产生1. 自然产生:由单性生殖产生。
未受精的雌、雄配子,甚至助细胞、反细胞等直接发育形成单倍体胚。
部分动物,如膜翅目昆虫(蜂、蚊)和某些同翅目昆虫(白蚁)的雄性个体都是孤雌生殖形成的自然单倍体。
2. 人工获得单倍体
花药培养:花药离体培养诱导配子体(花粉或子房)发育形成单倍体植株
这是应用最为广泛、成功的人工方法
种间或属间远缘杂交
栽培大麦(Hordeum vudare, 2n=2x=14)与野生球茎大麦(H. bulbosus, 2n=2x=14)杂种胚发育过程中,两物种染色体的行为不协调可导致球茎大麦的染色体逐渐丢失(称为染色体消减现象),可获得大麦的单倍体植株花药培养获得单倍体染色体消减获得单倍体大麦(四)、单倍体在遗传育种研究的应用1.提高育种的选择效率、加速育种进程
基因成单,加倍,纯合体
2.作为良好的遗传研究材料
基因成单,每个基因都发挥作用
3.用以分析染色体组间同源关系
部分同源关系六、非整倍体及其应用非整倍体:染色体数不是染色体组的整倍数的个体
某个体的染色体数比该物种的正常合子染色体数(2n)多或少一个以至若干个染色体。
三体 2n+l=(n–l)Ⅱ+Ⅲ
超倍体 四体 2n+2=(n-1)Ⅱ+Ⅳ
双三体 2n+1+1=(n-2)Ⅱ+2Ⅲ
单体 2n-1=(n-1)Ⅱ+Ⅰ
亚倍体 缺体 2n-2=(n-1)Ⅱ
双单体 2n-1-1=(n-2)Ⅱ+ 2Ⅰ
六、非整倍体及其应用非整倍体的类型
超倍体:多一条或几条染色体,遗传组成不平衡
亚倍体:少一条或几条染色体,遗传物质缺失
非整倍体的形成
减数分裂不正常产生n+1或n-1配子,后代为非整倍体
植物有丝分裂不正常也能产生非整倍体后代
非整倍体的存在
超倍体:二倍体、同源多倍体、异源多倍体均可能
亚倍体:二倍体、同源多倍体、异源多倍体(一)、单体1.单体的特点
●动物:单体的存在往往是许多动物的种性,许多昆虫(蝗虫、蟋蟀)的雌性为 XX型(即2n),雄性为XO型(即2n–l)
●植物:不同植物的单体表现有所不同
二倍体的单体:一般生活力极低而且不育
异源多倍体的单体:具有一定的生活力和育性
普通烟草(2n=4x=TTSS=48)的单体系列
普通小麦(2n=6x=AABBDD=42)的单体系列
(一)、单体普通烟草(2n=4x=TTSS=48)具有24种单体。
●分别用A, B, C, …, V, W, Z字母表示24条染色体;
●24种单体的表示为:
2n-IA, 2n-IB, 2n-IC, …, 2n-IW, 2n-IZ。
●各种单体具有不同的性状变异,表现在:
花冠大小、花萼大小、蒴果大小等性状上。普通小麦的单体系列普通小麦(2n=6x=AABBDD=42)具有21种单体。
●普通小麦的按ABD染色体组及部分同源关系编号为:
A组:1A, 2A, 3A, …, 6A, 7A;
B组:1B, 2B, 3B, …, 6B, 7B;
D组:1D, 2D, 3D, …, 6D, 7D。
●21种单体对应的表示方法为:
2n-I1A, 2n-I2A, …
2n-I1B, 2n-I2B, …
2n-I1D, 2n-I2D, …2.单体染色体的传递普通小麦单体:中期普通小麦单体:终变期硬粒小麦(2n=4x=AABB=28)1A单体2.单体染色体的传递普通小麦单体:四分体普通小麦单体:第一后期普通小麦单体染色体的传递单体自交后代 ●理论上:
◆配子 n: (n-1)=1:1
◆双:单:缺= 1:2:1
●影响因素:
◆单体被遗弃
◆ n, (n-1)参入受精不同
◆ 2n-I, 2n-II 成活率不同
导致不符合理论比例
如
(二)、缺体缺体一般都通过单体自交产生
在异源多倍体生物中可以存在
◆由于缺失一对染色体,对生物个体的性状表现的影响更大,生活力更差
◆普通烟草的缺体在幼胚阶段即死亡
◆普通小麦21种缺体都能够生存
遗传效应
◆表现广泛的性状变异(例)
◆通过缺体的性状变异,可能确定位于该染色体上的基因普通小麦缺体系列的穗形(三)、三体1.三体的性状变异
不同物种,不同染色体的三体的变异性状及程度不同
◆直果曼陀罗(2n=2x=24)的果型变异(图)
◆玉米(2n=2x=20)有10个不同的三体
◆普通小麦(2n=2x=42)具有21个三体,但性状变异较小
◆人类: XXX, XXY XYY三体的染色体联会与分离◆三体染色体联会与分离(图1、2、3、4、5、6)
(1)Ⅲ 2/1
(2)Ⅱ+Ⅰ 2/1 (单价体随机进入一个二分体细胞)
1/1 (单价体丢失)
◆形成四分体细胞:
(1)种类:n, n+1
(2)比例:n>n+1
◆配子育性与受精结合
配子育性:n>n+1, 尤其是在花粉中,因此n+1配子主要通过雌配子传递
后代: 小麦 双体(54.1%),三体(45%),四体(1%)
玉米 ♂ 三体1.4% 双体98.6%
♀ 三体24.5% 双体75.5%三体终变期:链式三价体三体染色体联会三体后期 I:2/1式分离三体末期 I:落后三价体复式三体的基因分离a 染色体随机分离(AAa)
(n+1) n
1,2/3 → AA AA a a
AAa → 1,3/2 → Aa Aa A A
2,3/1 → Aa Aa A A
AA:Aa:A:a=1:2:2:1
若n+l配子与n配子同等可育,且精子和卵子也同等可育,则自交子代的表现型比例就应该是[A]:[a]=35:1复式三体的基因分离假定复式三体的n+l精子全部不能参与受精,参与受精的卵子中1/4是n+l,3/4是n,则自交:
♀ ♂ 2A 1a
1AA
2Aa [A]:[a]=11:1
6A
3a复式三体染色体随机分离复式三体染色单体随机分离 ◆配子
n配子: A:4;a:2 (10:5)
n+1配子:AA:C42 = 6;
Aa: C41 C21 = 8;
aa: C21 = 1 (14:1)
配子:A:a = 24:1
◆表型
母本:3/4 (n+1)配子受精;
父本 (n+1)配子不授精;
表型:[A]:[a] = 10.3:1
b 染色单体随机分离(AAa)
分离的单位为6个染色单体:4A,2a
三价体的分离主要是2/1式,形成(n+1)和n
4!
则 AA配子=C24= = 6
(4-2)!2!
4! 2!
Aa配子=C14?C12= ? = 8
(4-1)!1! (2-1)!1!
2!
aa配子=C22= = 1
(2-2)!2!
即 AA : Aa : aa = 6:8:1
n配子与n+1配子数相等,即1(n+1):1n
n配子中 A:a=C14:C12=2:1=10:5
则全部配子种类和比例:
6AA:8Aa:1aa:10A:5a
若n+1配子与n配子同等可育,♀♂同等
参与受精,则自交子代:
(6AA:8Aa:1aa:10A:5a)2 = 24[A]:1[a]若参与受精的♀中3/4是n,1/4是n+1
n+1精子全部不能参与受精
则自交子代:
(6AA:8Aa:1aa:30A:15a)(2A:1a)
= 10.25[A]:1[a]
若同时考虑(n-1)Ⅱ中某一对基因如Bb
则自交子代:
(3B:1b)(10.25A:1a)(四)、四体与同源四倍体相比
◆只有一个同源组具有四条染色体
◆后期 I 2/2式分离的比例更高
四体小麦自交:四体73.8%, 三体23.6%, 双体1.9%
◆基因的分离与同源四倍体类似
◆生活力和配子的育性均更高(五)、非整倍体的应用1.基因的染色体定位(1).利用单体进行基因定位
隐性基因定位
普通烟草黄绿突变(yg2)的定位(过程)
单体定位机理
a表现双体与对应单体杂交(图)
a表现双体与非对应单体杂交(图)
显性基因定位
基本过程(p144)普通烟草黄绿突变(yg2)的定位普通烟草的黄绿突变型基因(隐性yg2)定位:
绿叶单体(2n-Ix)(共24种)×黄绿色叶型双体(yg2yg2)
↓
杂种F1(共24种)
(考察F1性状表现)
23种F1均表现为绿叶 仅与[2n-Is]杂种F1表现为:
绿叶和黄绿叶两种
(检查F1各个体的染色体数目)
所有绿叶个体均为双体(2n) yg2基因位于S染色体上?黄绿叶个体均为单体(2n-1)当隐性基因(a)在某染色体上时,则
P ((n–1)Ⅱ+ⅠA) × ((n–1)Ⅱ+Ⅱaa)
↙ ↘ ↓
n n–1 n
G (n–1) I+IA (n–1)I (n–1)I+Ia
↓ ↓
2n 2n–1
F1 ((n–1)Ⅱ+ⅡAa) ((n–1) Ⅱ+Ia)
A表现型 a表现型
双体 单体当隐性基因(a)不在某染色体上时,则
P ((n–1)ⅡAA +Ⅰ) × ((n–1)Ⅱaa +Ⅱ)
↙ ↘ ↓
n n–1 n
G (n–1) IA+I (n–1)IA (n–1)Ia+I
↓ ↓
2n 2n–1
F1 ((n–1)ⅡAa +Ⅱ) ((n–1) ⅡAa +I) 双体单体A表现型 A表现型a表现双体与对应单体杂交a表现双体与非对应单体杂交②单体测验显性基因
若显性基因(A)在某染色体上时,则F1单体自交后代中只有缺体是a表型
P ((n–1)Ⅱ+Ⅰa) × ((n–1)Ⅱ +ⅡAA )
↓
F1 ((n–1)Ⅱ+ⅡAa) ((n–1)Ⅱ+IA)
↓
♀ ♂ (n-1)Ⅰ+ IA (n-1) Ⅰ
F2 (n–1)Ⅱ+ⅡAA(n–1)Ⅱ+IA(n–1)Ⅱ+IA(n–1)Ⅱ 表现[a](n-1)Ⅰ+ IA(n-1) Ⅰ?当显性基因(A)不在某染色体上时,则其F1单体所自交的F2群体内,双体、单体、缺体植株都会有少数是a表现型的。
显性基因的单体定位过程隐性单体系列(n种)×显性纯合双体(AA)
↓
杂种F1(均表现为显性)(n种)
(进行染色体数目鉴定)
(单体↓自交)
F2(n种)
(鉴定性状表现、鉴定隐性F2的染色体数目)
n-1种隐性F2含双体、单体、缺体各种类型
只有1种F2双体、单体均为显性?基因所在1.基因的染色体定位(2).利用三体进行隐性基因定位
双体
自交后代表现型比例=3显:1隐
测交后代表现型比例=1显:1隐
三体
自交后代表现型比例≠3显:1隐
测交后代表现型比例≠1显:1隐② 三体测验
AAA×aa
↓
F1 AAa
↓
F2 所要测验的基因不在三体染色体上,
那么F2应该出现3A:1a
所要测验的基因在三体染色体上,
那么F2应该出现35A:1a或其他比例 ?(2)有目标地替换染色体
用单体有目标地替换染色体
已知某抗病基因R在小麦的6B染色体上,某优良品种不抗病(6BIIrr),要替换成抗病品种的6BIIRR:
(20II+6BIr) × (20II+6BIIRR)
?
F1 (20II+6BIIRr) (20II+6BIR)
?
F2 (20II+6BIIRR) 用单体并通过倍半二倍体有目标地替换染色体
使小麦只换取黑麦的个别染色体:
AABBDDRR × AABBDD
?
F1 AABBDDR
?
F2 (AABBDD+IR) × (AABBDD-IT)
?
20II+IR+IT
?
2n=20IITT+IIRR=21II用缺体有目标地替换染色体
已知某抗病基因R在小麦的6D染色体上,某优良品种不抗病(6DIIrr),要替换成抗病品种的6BIIRR:
(21II-6DII) × (20II+6DIIRR)
?
F1 (20II + 6DIR)
?
F2 (20II + 6DIIRR) 2.有目标地替换染色体小麦抗病品种: 20II+6BIIRR;
不抗病6B单体: 20II+ 6BIIrr;
20II+ 6BIr;x 20II+6BIIRR
20II+6BIIRr(双体抗病) 20II+6BIR (单体抗病)
?
20II+6BIIRR
小麦:
AABBDDRR x AABBDD
AABBDD+R(7I)
(2n+1): AABBDD+I1R
AABBDD+I2R
…….…
ABD+IR x ABD-IT
20II+IT+IR
?
(异代换系) 20IIT+IIR
异附加系:
AABBDD+II1R
……………
缺体
21II – 6DII x 20II+6DIIrr
20II+6DIR
?
20II+6DIIRR
?
同课章节目录
- 第一章 遗传因子的发现
- 第1节 盂德尔的豌豆杂交实验(一)
- 第2节 孟德尔的豌豆杂交实验(二)
- 第二章 基因和染色体的关系
- 第1节 减数分裂和受精作用
- 第2节 基因在染色体上
- 第3节 伴性遗传
- 第三章 基因的本质
- 第1节 DNA是主要的遗传物质
- 第2节 DNA分子的结构
- 第3节 DNA的复制
- 第4节 基因是有遗传效应的DNA片段
- 第四章 基因的表达
- 第1节 基因指导蛋白质的合成
- 第2节 基因对性状的控制
- 第3节 遗传密码的破译(选学)
- 第五章 基因突变及其他变异
- 第1节 基因突变和基因重组
- 第2节 染色体变异
- 第3节 人类遗传病
- 第六章 从杂交育种到基因工程
- 第1节 杂交育种与诱变育种
- 第2节 基因工程及其应用
- 第七章 现代生物进化理论
- 第1节 现代生物进化理论的由来
- 第2节 现代生物进化理论的主要内容