高中生物总复习讲解课件:专题6 光合作用(共41张PPT)
文档属性
| 名称 | 高中生物总复习讲解课件:专题6 光合作用(共41张PPT) |
|
|
| 格式 | pptx | ||
| 文件大小 | 2.0MB | ||
| 资源类型 | 试卷 | ||
| 版本资源 | 通用版 | ||
| 科目 | 生物学 | ||
| 更新时间 | 2024-09-23 00:00:00 | ||
图片预览




文档简介
(共41张PPT)
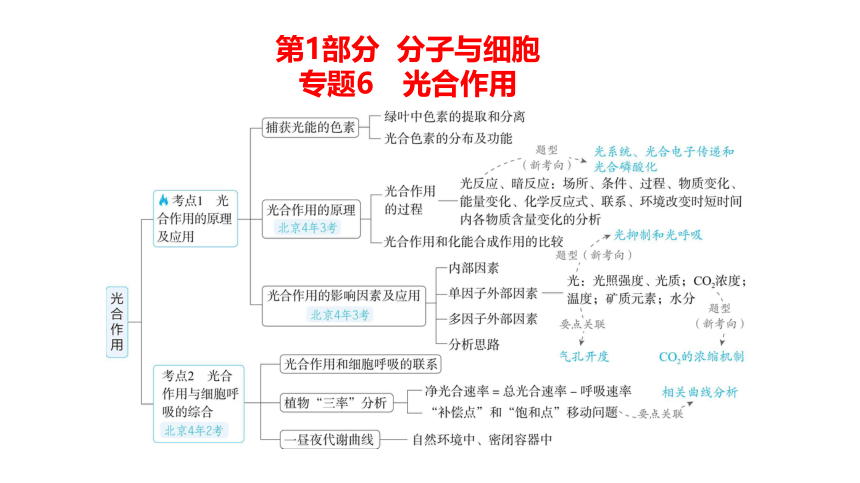
第1部分 分子与细胞
专题6 光合作用
考点1 光合作用的原理及应用
一、捕获光能的色素
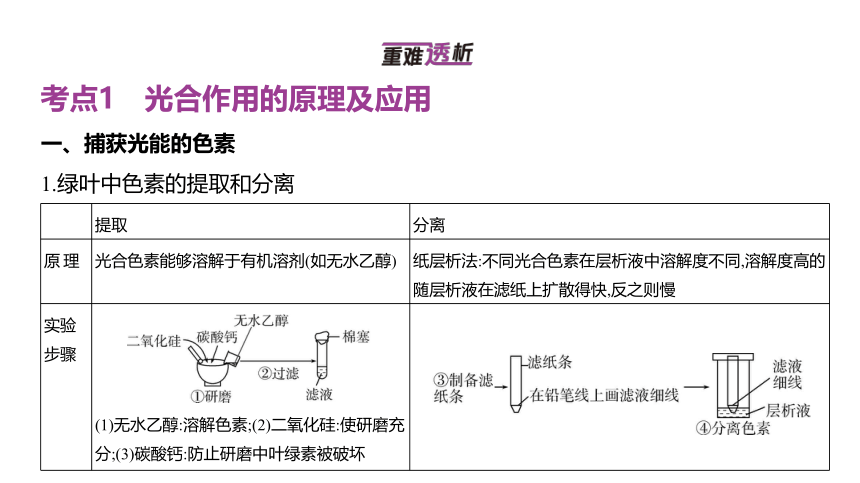
1.绿叶中色素的提取和分离
提取 分离
原理 光合色素能够溶解于有机溶剂(如无水乙醇) 纸层析法:不同光合色素在层析液中溶解度不同,溶解度高的随层析液在滤纸上扩散得快,反之则慢
实验 步骤 (1)无水乙醇:溶解色素;(2)二氧化硅:使研磨充分;(3)碳酸钙:防止研磨中叶绿素被破坏
实 验 结 果
(1)色素带的条数与色素种类有关,四条色素带说明有四种色素;(2)色素带的宽窄与色素含量有关,色素带越宽说明此种色素含量越多;(3)色素带扩散速度与溶解度有关,扩散速度越快说明溶解度越高
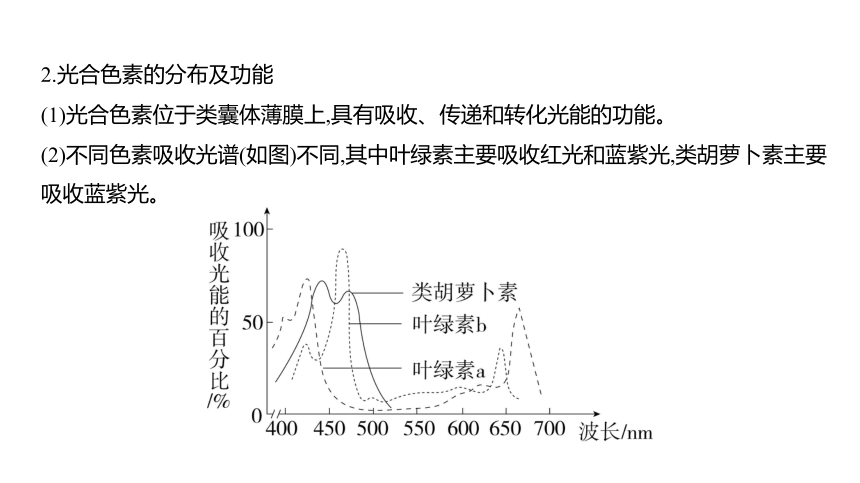
2.光合色素的分布及功能
(1)光合色素位于类囊体薄膜上,具有吸收、传递和转化光能的功能。
(2)不同色素吸收光谱(如图)不同,其中叶绿素主要吸收红光和蓝紫光,类胡萝卜素主要
吸收蓝紫光。
易混易错 (1)叶绿体中的色素(即光合色素)≠液泡中的色素(如水溶性的花青素)≠
光敏色素,光敏色素是一种接收光信号的分子,调节植物的生长发育。
(2)叶绿素的组成元素为C、H、O、N、Mg,类胡萝卜素的组成元素仅有C、H、O,故
土壤中缺少Mg可以使植株叶片黄化。
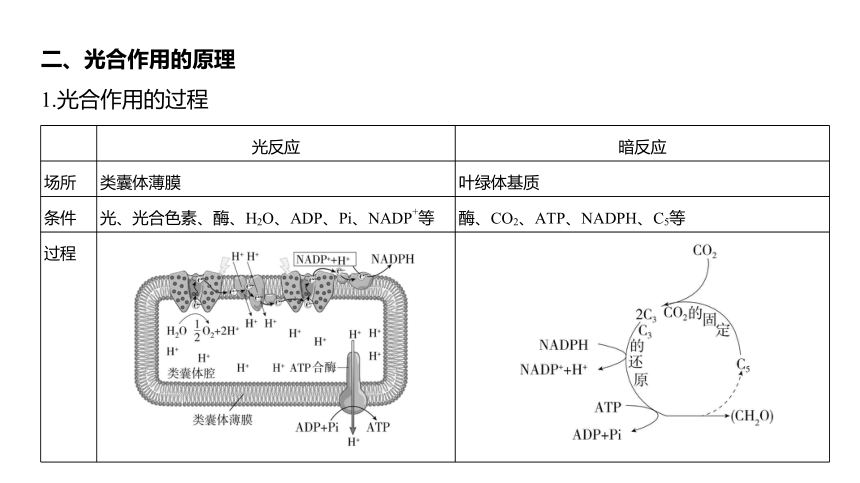
二、光合作用的原理
1.光合作用的过程
光反应 暗反应
场所 类囊体薄膜 叶绿体基质
条件 光、光合色素、酶、H2O、ADP、Pi、NADP+等 酶、CO2、ATP、NADPH、C5等
过程
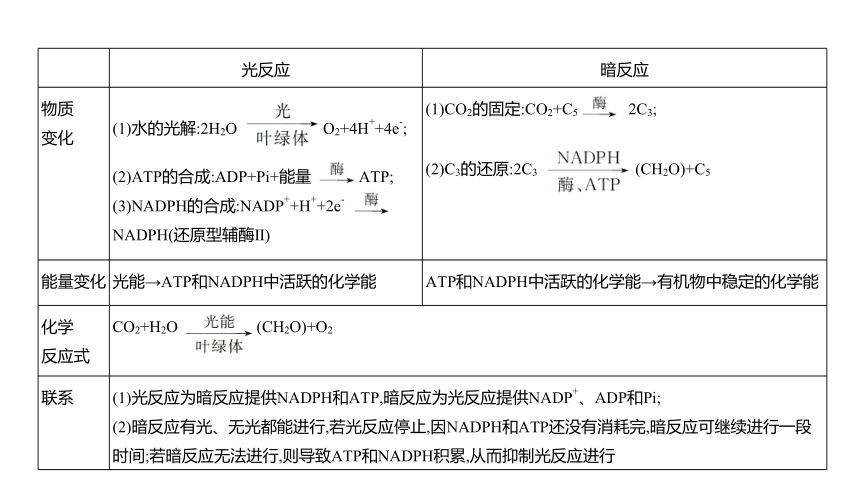
光反应 暗反应
物质 变化 (1)水的光解:2H2O O2+4H++4e-; (2)ATP的合成:ADP+Pi+能量 ATP; (3)NADPH的合成:NADP++H++2e- NADPH(还原型辅酶Ⅱ) (1)CO2的固定:CO2+C5 2C3;
(2)C3的还原:2C3 (CH2O)+C5
能量变化 光能→ATP和NADPH中活跃的化学能 ATP和NADPH中活跃的化学能→有机物中稳定的化学能
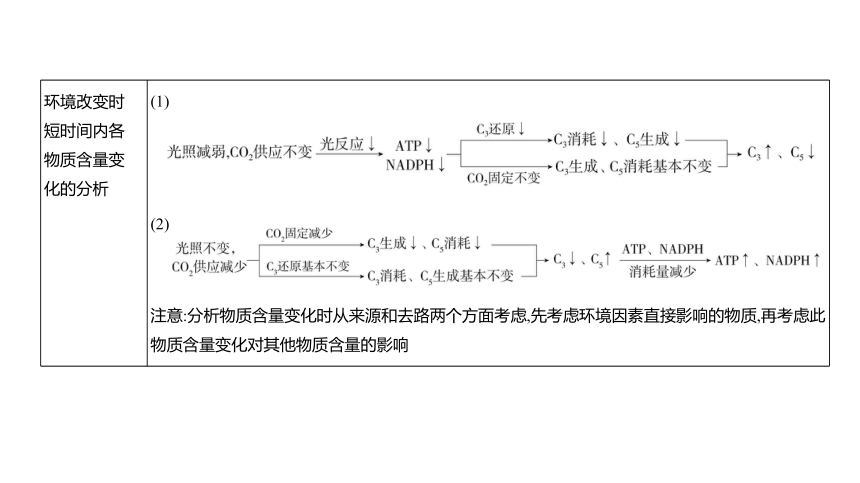
化学 反应式 CO2+H2O (CH2O)+O2 联系 (1)光反应为暗反应提供NADPH和ATP,暗反应为光反应提供NADP+、ADP和Pi; (2)暗反应有光、无光都能进行,若光反应停止,因NADPH和ATP还没有消耗完,暗反应可继续进行一段时间;若暗反应无法进行,则导致ATP和NADPH积累,从而抑制光反应进行 环境改变时 短时间内各 物质含量变 化的分析 (1)
(2)
注意:分析物质含量变化时从来源和去路两个方面考虑,先考虑环境因素直接影响的物质,再考虑此物质含量变化对其他物质含量的影响
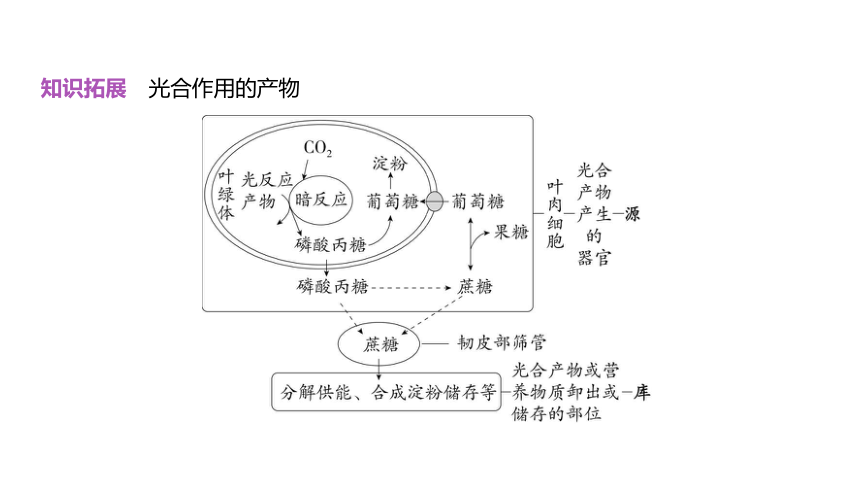
知识拓展 光合作用的产物
光合作用的产物主要是蔗糖、淀粉等糖类,蔗糖在细胞质基质中合成,淀粉在叶
绿体基质中合成,蔗糖最终经过韧皮部(筛管)由叶片(源)运往其他器官(库)。由于蔗
糖具有较高的水溶性和运输速率且性质稳定,因此蔗糖是糖类物质在植物体内运输的主
要形式。
小表达 将分离得到的叶绿体悬浮在适宜溶液中,照光后有氧气释放。如果在该适宜
溶液中将叶绿体的双层膜破裂后再照光, (填“有”或“没有”)氧气释放,原因是
。
有
类囊体膜是H2O分解释放O2的场所,叶绿体的双层膜破裂不影响类囊体膜的功能
光合作用 化能合成作用
区别 能量来源 光能 无机物氧化分解释放的化学能
代表生物 绿色植物 硝化细菌
相同 都能将CO2和H2O等无机物合成有机物 2.光合作用和化能合成作用的比较
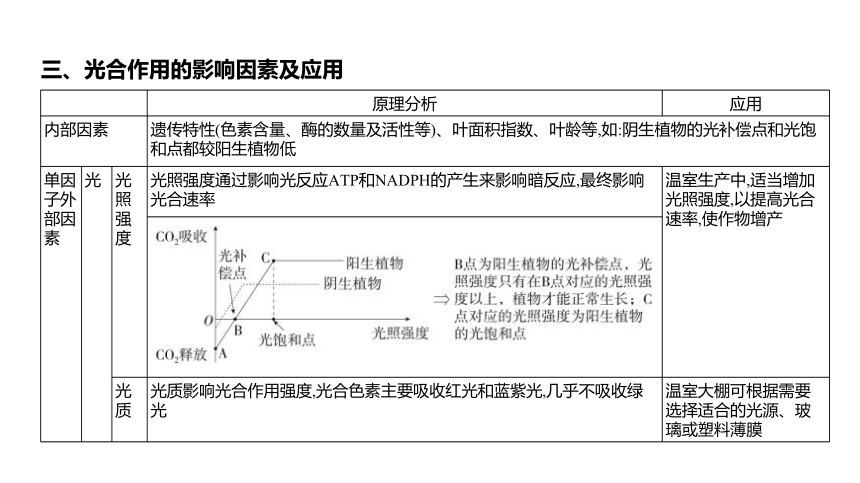
三、光合作用的影响因素及应用
原理分析 应用
内部因素 遗传特性(色素含量、酶的数量及活性等)、叶面积指数、叶龄等,如:阴生植物的光补偿点和光饱和点都较阳生植物低 单因 子外 部因 素 光 光 照 强 度 光照强度通过影响光反应ATP和NADPH的产生来影响暗反应,最终影响光合速率 温室生产中,适当增加光照强度,以提高光合速率,使作物增产
光 质 光质影响光合作用强度,光合色素主要吸收红光和蓝紫光,几乎不吸收绿光 温室大棚可根据需要选择适合的光源、玻璃或塑料薄膜
原理分析 应用
单 因 子 外 部 因 素 CO2浓度 农田“正其行、通其风”及温室大棚增施有机肥(微生物分解产生CO2)等均可增大CO2浓度,提高光合速率
温度 温室大棚内增加昼夜温差,有助于植物有机物的积累
矿质元素 (1)N、Mg等是合成叶绿素所必需的矿质元素; (2)P参与ATP的形成 根据作物的需肥规律适时、适量施肥,提高农作物产量
水分 (1)水是光合作用的原料; (2)缺水时,气孔开度减小,CO2吸收量减小 根据作物的需水规律合理灌溉
原理分析 应用
多因子 外部因素
(1)A点前:限制因子主要为光照强度。 (2)B点后:光照强度不再是限制因子,光合速率还受温度、CO2浓度等的影响 温室栽培时,在一定光照强度下,白天适当提高温度(也可同时适当增加CO2浓度)可提高光合速率;当温度适宜时,可适当增加光照强度和CO2浓度以提高光合速率
分 析 思 路 思路 写出光合作用的相关反应式→结合反应物、生成物、反应条件进行分析
实例 如图为CO2浓度对光合速率的影响曲线,试分析B
点前,光合速率随CO2浓度增加而增大的原因。B
点后,CO2浓度增加而光合速率不再增加,试分析此
时限制光合速率的因素可能有哪些
思路 分析 第一步:写出CO2所涉及的光合作用过程:CO2+C5 2C3 (CH2O)+C5。
第二步:分析。
B点前,光合速率随CO2浓度增加而增大的原因:CO2↑→C3↑→(CH2O)↑→光合速率↑。
B点后,CO2浓度增加,而光合速率不再增加,此时限制光合速率的因素有①反应物:C5含量不足;
②反应条件:酶活性不够高或数量不足,ATP、NADPH的含量较低;③生成物:(CH2O)积累
小表达 (1)大棚生产中,为什么多施有机肥能够提高作物的产量
。
(2)影响气孔开闭的因素有 ,气孔的开闭
(填“是”或“不是”)受单一因素的影响,举例说明: 。
干旱会影响脱落酸(ABA)的合成,从而影响植物气孔开闭
有机肥中的有机物被土壤中的微生物分解成了CO2和无机盐,CO2是光合作用的原料,无机盐有利于植物生长
光照、含水量、植物激素、温度等
不是
考点2 光合作用与细胞呼吸的综合
光合 作用 和细 胞呼 吸的 联系 物质 转变
能量 转化
植物 “三率” 分析
净光合速率=总光合速率-呼吸速率 净光合速率 总光合速率 呼吸速率
参照主体 细胞或植物体 叶绿体 线粒体(只考虑有氧呼吸)
CO2 吸收量 固定量、消耗量、利用量 释放量(黑暗)
O2 释放量 产生量 吸收量(黑暗)
有机物 积累量 制造量、合成量 消耗量(黑暗)
“补偿点”和“饱和点”移动问题:分析时可利用“光合条件若变好,补偿饱和两边跑;光合条件若变差,补偿饱和中间靠”来判断,如:光合作用最适温度25 ℃,细胞呼吸最适温度30 ℃,若温度由30 ℃降至25 ℃,图
中A、B、C三点分别向上(细胞呼吸速率减慢)、左(光合作用变强,若要光合作用等于呼吸作用,光照需变 弱)、右(光合作用变强,可固定更多光能) 一昼夜代谢曲线
自然环境中: 图1 密闭容器中:
图2
图1中AB段及图2中FG段可见,在夏季午时左右,光照强、温度高,叶片部分气孔关闭,CO2吸收速率降低,导致光合速率降低(即光合午休) 题型1 光系统、光合电子传递和光合磷酸化(新考向)
1.光系统:叶绿体进行能量转化需要依靠光系统,光系统是主要由光合色素和蛋白质结
合形成的大型复合物,包括光系统Ⅰ(PSⅠ)和光系统Ⅱ(PSⅡ),光系统能将光能转化为
电能(2020北京,19,12分)(2023湖北,8,2分)(2023山东,21,10分)。
2.光合电子传递和光合磷酸化:光系统产生的高能电子沿光合电子传递链依次传递,促
使NADPH的形成,同时建立了H+浓度梯度。光合磷酸化即H+顺浓度梯度驱动ATP合
酶的运转,将ADP磷酸化为ATP的过程,该过程与有氧呼吸第三阶段类似,如图所示。
(2023湖北,8,2分)(2022重庆,23,14分)
知识归纳 (1)光系统(PSⅠ、PSⅡ)分布在类囊体薄膜上,PSⅡ能进行水的光解,产生
O2、H+和电子(e-);PSⅠ能介导NADPH的产生。光反应中,电子的供体是水,电子的最终
受体是NADP+。
(2)H+进入类囊体腔的运输方式是主动运输,由高能电子供能;H+通过ATP合酶出类囊
体腔的运输方式是协助扩散。
典例 (2023通州期末,4)下图为高等植物叶绿体部分结构示意图,PSⅡ和PSⅠ系统是
由蛋白质和光合色素组成的复合物,下列相关说法错误的是 ( )
A.PSⅡ和PSⅠ系统分布在高等
植物叶绿体内膜上
B.H+向膜外转运过程释放的
能量为合成ATP供能
C.PSⅡ和PSⅠ系统中的光合色
素属于脂溶性物质
D.如果e-和H+不能正常传递给NADP+,暗反应的速率会下降
A
解析 PSⅡ和PSⅠ系统是由蛋白质和光合色素组成的复合物,光合色素存在于类囊
体薄膜上,A错误;H+向膜外转运过程中,电化学势能转化为ATP中活跃的化学能,为
ATP的合成提供能量,B正确;如果e-和H+不能正常传递给NADP+,会使NADPH的合成减
少,导致暗反应中C3的还原速率下降,D正确。
题型2 光抑制和光呼吸(新考向)
光抑制 定义 强光抑制光合作用的现象
产生机制 及危害 当光照强度超过叶绿体所能利用的最大光能时,过量光能会引起叶肉细胞内NADP+不足(光合色素过量吸收光能产生的电子会超过NADP+的可利用量),此时过量的电子导致光有毒物质(如活性氧)的产生。这些光有毒物质会攻击PSⅡ中的D1蛋白、膜结构、核酸等物质,损坏光合结构,使光合速率下降(2020北京,19,12分)(2023山东,21,10分)
光呼吸 (2023湖南, 17,12分) (2022江苏, 20,9分) 光呼吸与 暗反应的 关系
生理过程 光呼吸与植物体内一种双功能酶——Rubisco有关,当叶肉细胞中CO2浓度高时,该酶可催化C5固定CO2,进而合成有机物;当CO2浓度低时,该酶可催化C5与O2反应,产生C3、磷酸乙醇酸,磷酸乙醇酸经一系列反应产生CO2
生理意义 (1)光呼吸是进行光合作用的细胞为适应高光照及高O2、低CO2的条件下,提高抗逆性而形成的一条代谢途径;
(2)在干旱和高辐射等的环境中,气孔关闭,胞间CO2浓度降低,会导致光抑制。此时光呼吸释放CO2,消耗多余能量,对光合器官起保护作用,避免产生光抑制
典例 (2024西城期末,17)植物的光呼吸是在光下消耗氧气并释放CO2的过程,会导致
光合作用减弱、作物减产。研究人员为获得光诱导型高产水稻,在其叶绿体内构建一
条光呼吸支路(GMA途径)。
(1)图1所示光呼吸过程中,O2与CO2竞争结合 ,抑制了光合作用中的
阶段。同时乙醇酸从叶绿体进入过氧化物酶体在G酶的参与下进行代谢,
造成碳流失进而导致水稻减产。
(2)研究人员将外源G酶、A酶和M酶的基因导入水
稻细胞,使其在光诱导下表达,并在叶绿体中发挥作
用。检测发现,转基因水稻的净光合速率、植株干
重等方面均高于对照组。可利用图2所示模型解释
其原因,但图中存在两处错误,请圈出并改正。
R酶
暗反应/
CO2的固定
“A酶”改为“G酶”/“A酶”改为“G酶、M酶、A酶”;GMA途径使得CO2/O2↑
(3)研究人员测定了转基因水稻叶片中外源G酶基因的表达量,以及G酶总表达量随时
间的变化情况(图3)。
①外源G酶基因表达量与PFD(代表光合有效光辐射强度)大致呈正相关,仅在14时明
显下降,由此推测外源G酶基因表达除光强外,还可能受 等因
素的影响。
②据图3可知,12~14时 ,推测此时段转基因水稻光呼吸增强。
(4)茎中光合产物的堆积会降低水稻结实率而减产,而本研究中GMA途径的改造并未降
低水稻的结实率。结合上述研究将以下说法排序成合理解释:尽管GMA途径促进叶片
产生较多光合产物→ →水稻茎中有机物不至于过度堆积而保证结实率。
A.光呼吸增强使得光合产物未爆发式增加
B.光合产物可以及时运输到籽粒
C.G酶表达量的动态变化,使中午进入GMA途径的乙醇酸未显著增加
气孔导度/CO2浓度/温度
内源G酶表达量显著升高
C→A→B
解析 (1)结合图1分析,R酶既能结合CO2催化其固定(暗反应阶段),也能结合O2催化
生成2-PG(光呼吸),因此光呼吸过程中,O2与CO2竞争结合R酶,从而抑制了光合作用的
暗反应(或CO2的固定)。(2)结合图1信息:G酶催化乙醇酸转变为乙醛酸时消耗O2,降
低O2浓度;M酶催化乙醛酸生成苹果酸,苹果酸进一步转化为丙酮酸(此过程产生CO2),
丙酮酸进一步反应产生CO2,提高了细胞内的CO2浓度,上述过程导致CO2/O2↑,即图2中
GMA途径使得CO2/O2↑,乙醇酸→GMA途径中的“A酶”应改为“G酶”(或“G酶、M酶、
A酶”)。(3)①14时光照强度强、温度高,易导致气孔导度下降,CO2吸收减少,再结合
外源G酶基因表达量在14时明显下降,推测外源G酶基因表达除光强外,还可能与气孔
导度/CO2浓度/温度等因素有关。②根据图3可知,与其他时段相比,12~14时,转基因
水稻叶片中总G酶和外源G酶(在叶绿体中发挥作用)的表达量差值增大,说明内源G酶
(在过氧化物酶体中发挥作用)表达量显著升高,可反映过氧化物酶体中乙醇酸含量
升高,再结合图1中O2→2-PG→乙醇酸(转入过氧化物酶体中),推测此时段转基因水稻
光呼吸增强。(4)分析物质含量的变化时,可从该物质的来源、去路两个角度考虑。
水稻茎中有机物未发生过度堆积的原因:①有机物来源不会爆发式增加,原因为C选
项所述,即C→A;②有机物去路未被抑制或阻断,即B选项所述,故是C→A→B。
题型3 CO2的浓缩机制(新考向)
CO2浓度是限制光合作用的关键因素。因为Rubisco对CO2的亲和力较低,CO2浓度
过低时,其催化CO2固定的过程会受限制,并使光呼吸增强。所以,各类光合生物演化出
不同的机制来提高胞内CO2浓度,从而促进光合作用,降低光呼吸。
途径 主要内容 意义
蓝细菌羧化体途径 (2021天津,15,10分) 蓝细菌可通过主动运输( HC 进
入光合片层膜)和羧化体来提高
Rubisco周围的CO2浓度
途径 主要内容 意义
C4途径 CO2首先在叶肉细胞内被固定在四碳化合物(草酰乙酸,这也是“C4”名字的由来)中,草酰乙酸转化为苹果酸,然后被运输到维管束鞘细胞中脱羧,放出的CO2在Rubisco催化下再与C5反应生成C3(2023湖南,17, 12分) (1)PEP羧化酶对CO2的亲和力极高,提高了C4植物固定CO2的能力;
(2)PEP羧化酶起到“CO2泵”的作用,把CO2“压”进维管束细胞中,使Rubisco羧化部位的CO2浓度比
C3植物的高很多,从而减少光呼吸,因而C4植物在强光下具有比C3植物更高的光合速率;
(3)维管束鞘细胞中的光合产物可就近运入维管束,从而避免了光合产物累积对光合作用可能产生的抑制作用
途径 主要内容 意义
CAM途径 生活在干旱环境中的景天、仙人掌等植物夜间气孔开放,吸收CO2,经一系列反应形成苹果酸,储存在液泡中;白天气孔关闭,苹果酸转移到细胞质基质中脱羧,放出CO2,进入卡尔文循环 白天(高温),植物气孔关闭,避免了水分散失,有利于植物在干旱环境中生存
易混易错 (1)C3植物与C4植物在光反应阶段完全相同。卡尔文循环(又称C3途径)是
碳同化(即固定CO2,形成糖类)的基本途径,可合成糖类等产物,C4途径和CAM途径都只
起固定、运转CO2的作用,最终还是通过C3途径合成光合产物。
(2)CAM途径与C4途径基本相同,二者的主要差别在于C4植物中CO2固定和糖的合成是
在空间上(叶肉细胞和维管束鞘细胞)分开的,而CAM植物则是在时间上(黑夜和白天)
分开的。
典例1 (2024丰台一模,3)CAM植物白天气孔关闭,夜晚气孔打开,以适应干旱环境,如
图为其部分代谢途径,相关叙述不正确的是 ( )
A.催化过程①和过程②所需的酶不同
B.卡尔文循环的场所是叶绿体类囊体薄膜
C.CAM植物白天气孔关闭可减少水分散失
D.夜晚缺乏NADPH和ATP不能进行卡尔文循环
B
解析 酶具有专一性,一种酶只能催化一种或一类化学反应,过程①和②是两个不同
的过程,因此需要的酶不同,A正确。卡尔文循环是光合作用的暗反应过程,发生在叶
绿体基质中,B错误。CAM植物白天气孔关闭有利于减少蒸腾作用,减少水分的散失,C
正确。夜晚缺乏光照,CAM植物不能进行光反应,导致暗反应因缺乏NADPH和ATP而
无法进行,D正确。
典例2 (2021天津,15,10分)Rubisco是光合作用过程中催化CO2固定的酶。但其也能
催化O2与C5结合,形成C3和C2,导致光合效率下降。CO2与O2竞争性结合Rubisco的同一
活性位点,因此提高CO2浓度可以提高光合效率。
(1)蓝细菌具有CO2浓缩机制,如图所示。
据图分析,CO2依次以 和 方式通过细胞膜和光合片层膜。
蓝细菌的CO2浓缩机制可提高羧化体中Rubisco周围的CO2浓度,从而通过促进
和抑制 提高光合效率。
(2)向烟草内转入蓝细菌Rubisco的编码基因和羧化体外壳蛋白的编码基因。若蓝细菌
羧化体可在烟草中发挥作用并参与暗反应,应能利用电子显微镜在转基因烟草细胞的
中观察到羧化体。
(3)研究发现,转基因烟草的光合速率并未提高。若再转入HC 和CO2转运蛋白基因
并成功表达和发挥作用,理论上该转基因植株暗反应水平应 ,光反应水平应
,从而提高光合速率。
自由扩散
主动运输
CO2 固定
O2与C5结合
叶绿体
提高
提高
解析 (1)从蓝细菌的CO2浓缩机制图示中可以看出CO2穿过细胞膜为自由扩散,穿过
光合片层膜时需借助CO2转运蛋白并消耗能量,为主动运输。由题意可知,Rubisco既能
催化CO2固定,又能催化O2与C5结合,蓝细菌可通过CO2浓缩机制提高羧化体中Rubisco
周围的CO2浓度,从而通过促进 CO2固定和抑制O2与C5结合来提高光合效率。(2)烟草
细胞为真核细胞,进行暗反应的场所为叶绿体基质。若蓝细菌羧化体能在烟草中发挥
作用并参与暗反应,应能利用电子显微镜在转基因烟草细胞的叶绿体中观察到羧化
体。(3)根据题意可知,HC 和CO2转运蛋白有助于提高羧化体内CO2的浓度,从而提高
转基因植株的暗反应水平,暗反应水平提高可为光反应提供更多的NADP+和ADP等,
提高光反应水平,从而提高光合速率。
第1部分 分子与细胞
专题6 光合作用
考点1 光合作用的原理及应用
一、捕获光能的色素
1.绿叶中色素的提取和分离
提取 分离
原理 光合色素能够溶解于有机溶剂(如无水乙醇) 纸层析法:不同光合色素在层析液中溶解度不同,溶解度高的随层析液在滤纸上扩散得快,反之则慢
实验 步骤 (1)无水乙醇:溶解色素;(2)二氧化硅:使研磨充分;(3)碳酸钙:防止研磨中叶绿素被破坏
实 验 结 果
(1)色素带的条数与色素种类有关,四条色素带说明有四种色素;(2)色素带的宽窄与色素含量有关,色素带越宽说明此种色素含量越多;(3)色素带扩散速度与溶解度有关,扩散速度越快说明溶解度越高
2.光合色素的分布及功能
(1)光合色素位于类囊体薄膜上,具有吸收、传递和转化光能的功能。
(2)不同色素吸收光谱(如图)不同,其中叶绿素主要吸收红光和蓝紫光,类胡萝卜素主要
吸收蓝紫光。
易混易错 (1)叶绿体中的色素(即光合色素)≠液泡中的色素(如水溶性的花青素)≠
光敏色素,光敏色素是一种接收光信号的分子,调节植物的生长发育。
(2)叶绿素的组成元素为C、H、O、N、Mg,类胡萝卜素的组成元素仅有C、H、O,故
土壤中缺少Mg可以使植株叶片黄化。
二、光合作用的原理
1.光合作用的过程
光反应 暗反应
场所 类囊体薄膜 叶绿体基质
条件 光、光合色素、酶、H2O、ADP、Pi、NADP+等 酶、CO2、ATP、NADPH、C5等
过程
光反应 暗反应
物质 变化 (1)水的光解:2H2O O2+4H++4e-; (2)ATP的合成:ADP+Pi+能量 ATP; (3)NADPH的合成:NADP++H++2e- NADPH(还原型辅酶Ⅱ) (1)CO2的固定:CO2+C5 2C3;
(2)C3的还原:2C3 (CH2O)+C5
能量变化 光能→ATP和NADPH中活跃的化学能 ATP和NADPH中活跃的化学能→有机物中稳定的化学能
化学 反应式 CO2+H2O (CH2O)+O2 联系 (1)光反应为暗反应提供NADPH和ATP,暗反应为光反应提供NADP+、ADP和Pi; (2)暗反应有光、无光都能进行,若光反应停止,因NADPH和ATP还没有消耗完,暗反应可继续进行一段时间;若暗反应无法进行,则导致ATP和NADPH积累,从而抑制光反应进行 环境改变时 短时间内各 物质含量变 化的分析 (1)
(2)
注意:分析物质含量变化时从来源和去路两个方面考虑,先考虑环境因素直接影响的物质,再考虑此物质含量变化对其他物质含量的影响
知识拓展 光合作用的产物
光合作用的产物主要是蔗糖、淀粉等糖类,蔗糖在细胞质基质中合成,淀粉在叶
绿体基质中合成,蔗糖最终经过韧皮部(筛管)由叶片(源)运往其他器官(库)。由于蔗
糖具有较高的水溶性和运输速率且性质稳定,因此蔗糖是糖类物质在植物体内运输的主
要形式。
小表达 将分离得到的叶绿体悬浮在适宜溶液中,照光后有氧气释放。如果在该适宜
溶液中将叶绿体的双层膜破裂后再照光, (填“有”或“没有”)氧气释放,原因是
。
有
类囊体膜是H2O分解释放O2的场所,叶绿体的双层膜破裂不影响类囊体膜的功能
光合作用 化能合成作用
区别 能量来源 光能 无机物氧化分解释放的化学能
代表生物 绿色植物 硝化细菌
相同 都能将CO2和H2O等无机物合成有机物 2.光合作用和化能合成作用的比较
三、光合作用的影响因素及应用
原理分析 应用
内部因素 遗传特性(色素含量、酶的数量及活性等)、叶面积指数、叶龄等,如:阴生植物的光补偿点和光饱和点都较阳生植物低 单因 子外 部因 素 光 光 照 强 度 光照强度通过影响光反应ATP和NADPH的产生来影响暗反应,最终影响光合速率 温室生产中,适当增加光照强度,以提高光合速率,使作物增产
光 质 光质影响光合作用强度,光合色素主要吸收红光和蓝紫光,几乎不吸收绿光 温室大棚可根据需要选择适合的光源、玻璃或塑料薄膜
原理分析 应用
单 因 子 外 部 因 素 CO2浓度 农田“正其行、通其风”及温室大棚增施有机肥(微生物分解产生CO2)等均可增大CO2浓度,提高光合速率
温度 温室大棚内增加昼夜温差,有助于植物有机物的积累
矿质元素 (1)N、Mg等是合成叶绿素所必需的矿质元素; (2)P参与ATP的形成 根据作物的需肥规律适时、适量施肥,提高农作物产量
水分 (1)水是光合作用的原料; (2)缺水时,气孔开度减小,CO2吸收量减小 根据作物的需水规律合理灌溉
原理分析 应用
多因子 外部因素
(1)A点前:限制因子主要为光照强度。 (2)B点后:光照强度不再是限制因子,光合速率还受温度、CO2浓度等的影响 温室栽培时,在一定光照强度下,白天适当提高温度(也可同时适当增加CO2浓度)可提高光合速率;当温度适宜时,可适当增加光照强度和CO2浓度以提高光合速率
分 析 思 路 思路 写出光合作用的相关反应式→结合反应物、生成物、反应条件进行分析
实例 如图为CO2浓度对光合速率的影响曲线,试分析B
点前,光合速率随CO2浓度增加而增大的原因。B
点后,CO2浓度增加而光合速率不再增加,试分析此
时限制光合速率的因素可能有哪些
思路 分析 第一步:写出CO2所涉及的光合作用过程:CO2+C5 2C3 (CH2O)+C5。
第二步:分析。
B点前,光合速率随CO2浓度增加而增大的原因:CO2↑→C3↑→(CH2O)↑→光合速率↑。
B点后,CO2浓度增加,而光合速率不再增加,此时限制光合速率的因素有①反应物:C5含量不足;
②反应条件:酶活性不够高或数量不足,ATP、NADPH的含量较低;③生成物:(CH2O)积累
小表达 (1)大棚生产中,为什么多施有机肥能够提高作物的产量
。
(2)影响气孔开闭的因素有 ,气孔的开闭
(填“是”或“不是”)受单一因素的影响,举例说明: 。
干旱会影响脱落酸(ABA)的合成,从而影响植物气孔开闭
有机肥中的有机物被土壤中的微生物分解成了CO2和无机盐,CO2是光合作用的原料,无机盐有利于植物生长
光照、含水量、植物激素、温度等
不是
考点2 光合作用与细胞呼吸的综合
光合 作用 和细 胞呼 吸的 联系 物质 转变
能量 转化
植物 “三率” 分析
净光合速率=总光合速率-呼吸速率 净光合速率 总光合速率 呼吸速率
参照主体 细胞或植物体 叶绿体 线粒体(只考虑有氧呼吸)
CO2 吸收量 固定量、消耗量、利用量 释放量(黑暗)
O2 释放量 产生量 吸收量(黑暗)
有机物 积累量 制造量、合成量 消耗量(黑暗)
“补偿点”和“饱和点”移动问题:分析时可利用“光合条件若变好,补偿饱和两边跑;光合条件若变差,补偿饱和中间靠”来判断,如:光合作用最适温度25 ℃,细胞呼吸最适温度30 ℃,若温度由30 ℃降至25 ℃,图
中A、B、C三点分别向上(细胞呼吸速率减慢)、左(光合作用变强,若要光合作用等于呼吸作用,光照需变 弱)、右(光合作用变强,可固定更多光能) 一昼夜代谢曲线
自然环境中: 图1 密闭容器中:
图2
图1中AB段及图2中FG段可见,在夏季午时左右,光照强、温度高,叶片部分气孔关闭,CO2吸收速率降低,导致光合速率降低(即光合午休) 题型1 光系统、光合电子传递和光合磷酸化(新考向)
1.光系统:叶绿体进行能量转化需要依靠光系统,光系统是主要由光合色素和蛋白质结
合形成的大型复合物,包括光系统Ⅰ(PSⅠ)和光系统Ⅱ(PSⅡ),光系统能将光能转化为
电能(2020北京,19,12分)(2023湖北,8,2分)(2023山东,21,10分)。
2.光合电子传递和光合磷酸化:光系统产生的高能电子沿光合电子传递链依次传递,促
使NADPH的形成,同时建立了H+浓度梯度。光合磷酸化即H+顺浓度梯度驱动ATP合
酶的运转,将ADP磷酸化为ATP的过程,该过程与有氧呼吸第三阶段类似,如图所示。
(2023湖北,8,2分)(2022重庆,23,14分)
知识归纳 (1)光系统(PSⅠ、PSⅡ)分布在类囊体薄膜上,PSⅡ能进行水的光解,产生
O2、H+和电子(e-);PSⅠ能介导NADPH的产生。光反应中,电子的供体是水,电子的最终
受体是NADP+。
(2)H+进入类囊体腔的运输方式是主动运输,由高能电子供能;H+通过ATP合酶出类囊
体腔的运输方式是协助扩散。
典例 (2023通州期末,4)下图为高等植物叶绿体部分结构示意图,PSⅡ和PSⅠ系统是
由蛋白质和光合色素组成的复合物,下列相关说法错误的是 ( )
A.PSⅡ和PSⅠ系统分布在高等
植物叶绿体内膜上
B.H+向膜外转运过程释放的
能量为合成ATP供能
C.PSⅡ和PSⅠ系统中的光合色
素属于脂溶性物质
D.如果e-和H+不能正常传递给NADP+,暗反应的速率会下降
A
解析 PSⅡ和PSⅠ系统是由蛋白质和光合色素组成的复合物,光合色素存在于类囊
体薄膜上,A错误;H+向膜外转运过程中,电化学势能转化为ATP中活跃的化学能,为
ATP的合成提供能量,B正确;如果e-和H+不能正常传递给NADP+,会使NADPH的合成减
少,导致暗反应中C3的还原速率下降,D正确。
题型2 光抑制和光呼吸(新考向)
光抑制 定义 强光抑制光合作用的现象
产生机制 及危害 当光照强度超过叶绿体所能利用的最大光能时,过量光能会引起叶肉细胞内NADP+不足(光合色素过量吸收光能产生的电子会超过NADP+的可利用量),此时过量的电子导致光有毒物质(如活性氧)的产生。这些光有毒物质会攻击PSⅡ中的D1蛋白、膜结构、核酸等物质,损坏光合结构,使光合速率下降(2020北京,19,12分)(2023山东,21,10分)
光呼吸 (2023湖南, 17,12分) (2022江苏, 20,9分) 光呼吸与 暗反应的 关系
生理过程 光呼吸与植物体内一种双功能酶——Rubisco有关,当叶肉细胞中CO2浓度高时,该酶可催化C5固定CO2,进而合成有机物;当CO2浓度低时,该酶可催化C5与O2反应,产生C3、磷酸乙醇酸,磷酸乙醇酸经一系列反应产生CO2
生理意义 (1)光呼吸是进行光合作用的细胞为适应高光照及高O2、低CO2的条件下,提高抗逆性而形成的一条代谢途径;
(2)在干旱和高辐射等的环境中,气孔关闭,胞间CO2浓度降低,会导致光抑制。此时光呼吸释放CO2,消耗多余能量,对光合器官起保护作用,避免产生光抑制
典例 (2024西城期末,17)植物的光呼吸是在光下消耗氧气并释放CO2的过程,会导致
光合作用减弱、作物减产。研究人员为获得光诱导型高产水稻,在其叶绿体内构建一
条光呼吸支路(GMA途径)。
(1)图1所示光呼吸过程中,O2与CO2竞争结合 ,抑制了光合作用中的
阶段。同时乙醇酸从叶绿体进入过氧化物酶体在G酶的参与下进行代谢,
造成碳流失进而导致水稻减产。
(2)研究人员将外源G酶、A酶和M酶的基因导入水
稻细胞,使其在光诱导下表达,并在叶绿体中发挥作
用。检测发现,转基因水稻的净光合速率、植株干
重等方面均高于对照组。可利用图2所示模型解释
其原因,但图中存在两处错误,请圈出并改正。
R酶
暗反应/
CO2的固定
“A酶”改为“G酶”/“A酶”改为“G酶、M酶、A酶”;GMA途径使得CO2/O2↑
(3)研究人员测定了转基因水稻叶片中外源G酶基因的表达量,以及G酶总表达量随时
间的变化情况(图3)。
①外源G酶基因表达量与PFD(代表光合有效光辐射强度)大致呈正相关,仅在14时明
显下降,由此推测外源G酶基因表达除光强外,还可能受 等因
素的影响。
②据图3可知,12~14时 ,推测此时段转基因水稻光呼吸增强。
(4)茎中光合产物的堆积会降低水稻结实率而减产,而本研究中GMA途径的改造并未降
低水稻的结实率。结合上述研究将以下说法排序成合理解释:尽管GMA途径促进叶片
产生较多光合产物→ →水稻茎中有机物不至于过度堆积而保证结实率。
A.光呼吸增强使得光合产物未爆发式增加
B.光合产物可以及时运输到籽粒
C.G酶表达量的动态变化,使中午进入GMA途径的乙醇酸未显著增加
气孔导度/CO2浓度/温度
内源G酶表达量显著升高
C→A→B
解析 (1)结合图1分析,R酶既能结合CO2催化其固定(暗反应阶段),也能结合O2催化
生成2-PG(光呼吸),因此光呼吸过程中,O2与CO2竞争结合R酶,从而抑制了光合作用的
暗反应(或CO2的固定)。(2)结合图1信息:G酶催化乙醇酸转变为乙醛酸时消耗O2,降
低O2浓度;M酶催化乙醛酸生成苹果酸,苹果酸进一步转化为丙酮酸(此过程产生CO2),
丙酮酸进一步反应产生CO2,提高了细胞内的CO2浓度,上述过程导致CO2/O2↑,即图2中
GMA途径使得CO2/O2↑,乙醇酸→GMA途径中的“A酶”应改为“G酶”(或“G酶、M酶、
A酶”)。(3)①14时光照强度强、温度高,易导致气孔导度下降,CO2吸收减少,再结合
外源G酶基因表达量在14时明显下降,推测外源G酶基因表达除光强外,还可能与气孔
导度/CO2浓度/温度等因素有关。②根据图3可知,与其他时段相比,12~14时,转基因
水稻叶片中总G酶和外源G酶(在叶绿体中发挥作用)的表达量差值增大,说明内源G酶
(在过氧化物酶体中发挥作用)表达量显著升高,可反映过氧化物酶体中乙醇酸含量
升高,再结合图1中O2→2-PG→乙醇酸(转入过氧化物酶体中),推测此时段转基因水稻
光呼吸增强。(4)分析物质含量的变化时,可从该物质的来源、去路两个角度考虑。
水稻茎中有机物未发生过度堆积的原因:①有机物来源不会爆发式增加,原因为C选
项所述,即C→A;②有机物去路未被抑制或阻断,即B选项所述,故是C→A→B。
题型3 CO2的浓缩机制(新考向)
CO2浓度是限制光合作用的关键因素。因为Rubisco对CO2的亲和力较低,CO2浓度
过低时,其催化CO2固定的过程会受限制,并使光呼吸增强。所以,各类光合生物演化出
不同的机制来提高胞内CO2浓度,从而促进光合作用,降低光呼吸。
途径 主要内容 意义
蓝细菌羧化体途径 (2021天津,15,10分) 蓝细菌可通过主动运输( HC 进
入光合片层膜)和羧化体来提高
Rubisco周围的CO2浓度
途径 主要内容 意义
C4途径 CO2首先在叶肉细胞内被固定在四碳化合物(草酰乙酸,这也是“C4”名字的由来)中,草酰乙酸转化为苹果酸,然后被运输到维管束鞘细胞中脱羧,放出的CO2在Rubisco催化下再与C5反应生成C3(2023湖南,17, 12分) (1)PEP羧化酶对CO2的亲和力极高,提高了C4植物固定CO2的能力;
(2)PEP羧化酶起到“CO2泵”的作用,把CO2“压”进维管束细胞中,使Rubisco羧化部位的CO2浓度比
C3植物的高很多,从而减少光呼吸,因而C4植物在强光下具有比C3植物更高的光合速率;
(3)维管束鞘细胞中的光合产物可就近运入维管束,从而避免了光合产物累积对光合作用可能产生的抑制作用
途径 主要内容 意义
CAM途径 生活在干旱环境中的景天、仙人掌等植物夜间气孔开放,吸收CO2,经一系列反应形成苹果酸,储存在液泡中;白天气孔关闭,苹果酸转移到细胞质基质中脱羧,放出CO2,进入卡尔文循环 白天(高温),植物气孔关闭,避免了水分散失,有利于植物在干旱环境中生存
易混易错 (1)C3植物与C4植物在光反应阶段完全相同。卡尔文循环(又称C3途径)是
碳同化(即固定CO2,形成糖类)的基本途径,可合成糖类等产物,C4途径和CAM途径都只
起固定、运转CO2的作用,最终还是通过C3途径合成光合产物。
(2)CAM途径与C4途径基本相同,二者的主要差别在于C4植物中CO2固定和糖的合成是
在空间上(叶肉细胞和维管束鞘细胞)分开的,而CAM植物则是在时间上(黑夜和白天)
分开的。
典例1 (2024丰台一模,3)CAM植物白天气孔关闭,夜晚气孔打开,以适应干旱环境,如
图为其部分代谢途径,相关叙述不正确的是 ( )
A.催化过程①和过程②所需的酶不同
B.卡尔文循环的场所是叶绿体类囊体薄膜
C.CAM植物白天气孔关闭可减少水分散失
D.夜晚缺乏NADPH和ATP不能进行卡尔文循环
B
解析 酶具有专一性,一种酶只能催化一种或一类化学反应,过程①和②是两个不同
的过程,因此需要的酶不同,A正确。卡尔文循环是光合作用的暗反应过程,发生在叶
绿体基质中,B错误。CAM植物白天气孔关闭有利于减少蒸腾作用,减少水分的散失,C
正确。夜晚缺乏光照,CAM植物不能进行光反应,导致暗反应因缺乏NADPH和ATP而
无法进行,D正确。
典例2 (2021天津,15,10分)Rubisco是光合作用过程中催化CO2固定的酶。但其也能
催化O2与C5结合,形成C3和C2,导致光合效率下降。CO2与O2竞争性结合Rubisco的同一
活性位点,因此提高CO2浓度可以提高光合效率。
(1)蓝细菌具有CO2浓缩机制,如图所示。
据图分析,CO2依次以 和 方式通过细胞膜和光合片层膜。
蓝细菌的CO2浓缩机制可提高羧化体中Rubisco周围的CO2浓度,从而通过促进
和抑制 提高光合效率。
(2)向烟草内转入蓝细菌Rubisco的编码基因和羧化体外壳蛋白的编码基因。若蓝细菌
羧化体可在烟草中发挥作用并参与暗反应,应能利用电子显微镜在转基因烟草细胞的
中观察到羧化体。
(3)研究发现,转基因烟草的光合速率并未提高。若再转入HC 和CO2转运蛋白基因
并成功表达和发挥作用,理论上该转基因植株暗反应水平应 ,光反应水平应
,从而提高光合速率。
自由扩散
主动运输
CO2 固定
O2与C5结合
叶绿体
提高
提高
解析 (1)从蓝细菌的CO2浓缩机制图示中可以看出CO2穿过细胞膜为自由扩散,穿过
光合片层膜时需借助CO2转运蛋白并消耗能量,为主动运输。由题意可知,Rubisco既能
催化CO2固定,又能催化O2与C5结合,蓝细菌可通过CO2浓缩机制提高羧化体中Rubisco
周围的CO2浓度,从而通过促进 CO2固定和抑制O2与C5结合来提高光合效率。(2)烟草
细胞为真核细胞,进行暗反应的场所为叶绿体基质。若蓝细菌羧化体能在烟草中发挥
作用并参与暗反应,应能利用电子显微镜在转基因烟草细胞的叶绿体中观察到羧化
体。(3)根据题意可知,HC 和CO2转运蛋白有助于提高羧化体内CO2的浓度,从而提高
转基因植株的暗反应水平,暗反应水平提高可为光反应提供更多的NADP+和ADP等,
提高光反应水平,从而提高光合速率。
同课章节目录