2017六年级科学上册第一章第3课植物的光合作用课件新人教版
文档属性
| 名称 | 2017六年级科学上册第一章第3课植物的光合作用课件新人教版 |

|
|
| 格式 | zip | ||
| 文件大小 | 2.1MB | ||
| 资源类型 | 教案 | ||
| 版本资源 | 人教版(新课程标准) | ||
| 科目 | 科学 | ||
| 更新时间 | 2017-10-29 00:00:00 | ||
图片预览








文档简介
课件114张PPT。Chapter3 Photosynthesis in Plant本章内容:
光合作用的意义和研究历史
叶绿体和光合色素
光合作用机理
光呼吸
影响光合速率的外界因素
光合作用与农业生产
概述
一、自养植物和异养植物
1、异养植物(Heterophyte)
2、自养植物(Autophyte)
二、碳素同化作用(Carbon assimilation)
1、光合作用(Photosynthesis)
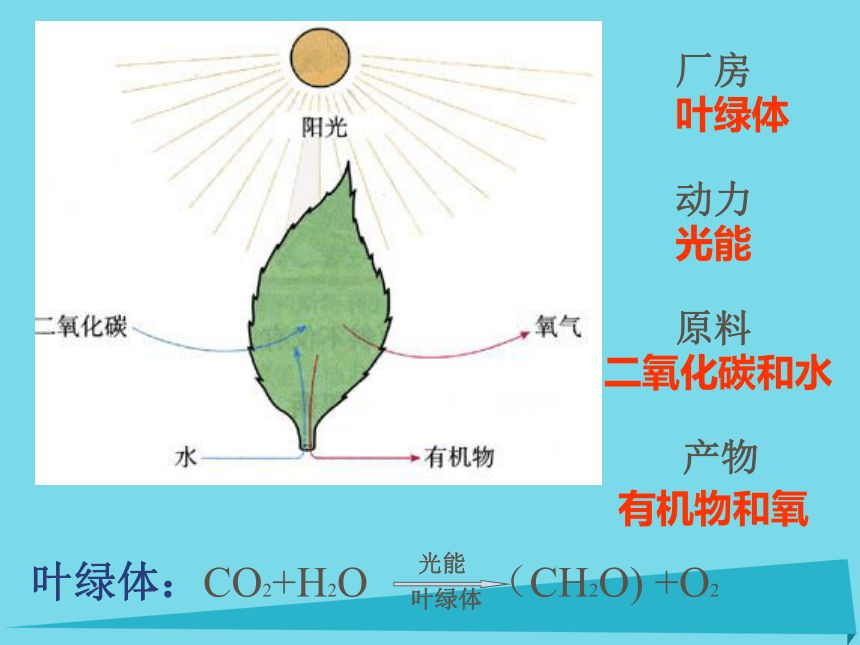
光
CO2 +H2O (CH2O ) + O2
叶绿体什么是光合作用? 绿色植物在光下,把二氧化碳和水转化为糖,并释放出氧气的过程。
厂房叶绿体动力光能原料二氧化碳和水产物有机物和氧 2、细菌光合作用 (Bacterial photosynthesis)
光、叶绿素
CO2 + H2S CH2O + H2O+S
3、化能合成作用(Chemosynthesis)
化能合成细菌
三、光合作用的重要性
1、有机物质的重要来源
2、把光能转化成化学能
3、大气中氧气的重要来源
第一节 叶绿体和叶绿体色素I一、叶绿体(Chloroplast)
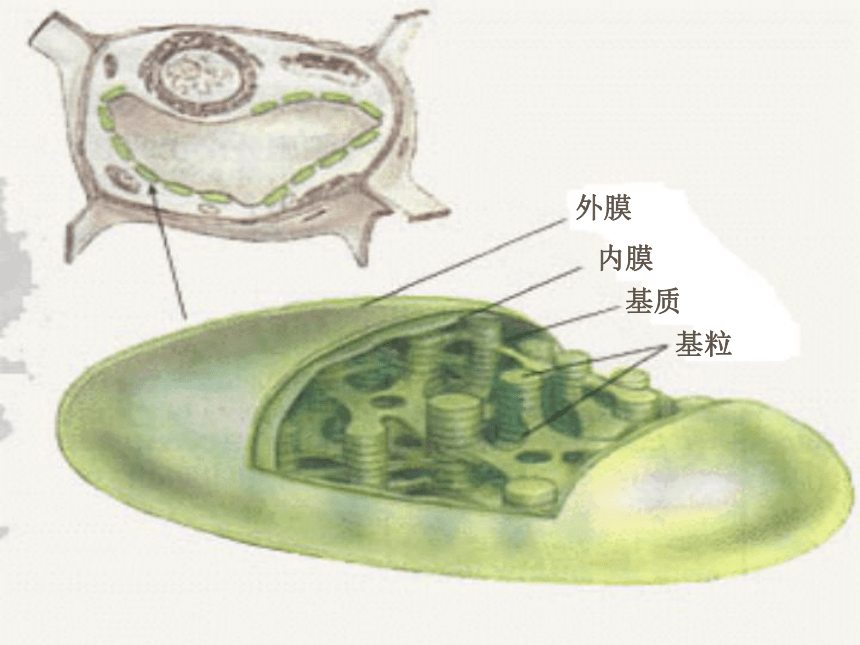
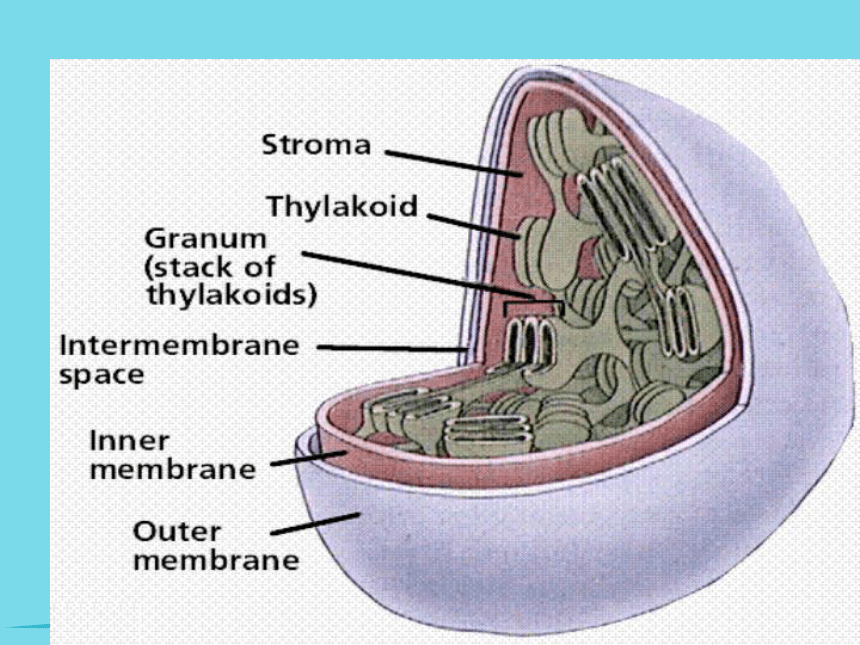
㈠叶绿体的结构:
椭圆形,一般直径为3~6um,厚为2~3um。每平方毫米的蓖麻叶就含3~5百万个叶绿体。
1、叶绿体膜—选择性屏障,控制物质进出。
2 、基质—CO2的固定,淀粉的合成和储藏(含酶类)
3 、基粒—光能-化学能(光合色素)
4 、嗜锇滴—基质中与锇酸容易结合的颗粒(醌类)
5 、类囊体—光合作用能量转换(又称光合膜)
基粒类囊体 (grana thylakoid)
基质类囊体 (stroma thylakoid)下一页外膜内膜基质基粒㈡叶绿体的成分
1、 水分(75%)
2 、 蛋白质(30~45%)—催化剂
3 、脂类(20~40%)—膜成分
4 、色素(8%)—与蛋白质结合,电子传递
5 、无机盐(10%)
6 、储藏物质(如淀粉等,10~20%)
7 、NAD+、NADP+、醌(如质体醌),起传递氢原子或电子的作用。二、光合色素
1、分类
叶绿素:类胡萝卜素=3:1 所以叶片一般呈绿色
叶绿素a:叶绿素b=3:1
叶黄素:胡萝卜素=2:1
解释:秋后或衰老的叶片多呈黄色,秋后枫树叶子呈红色2、光合色素化学结构与性质
⑴叶绿素(chlorophyll)
叶绿素不溶于水,但能溶于酒精、丙酮和石油醚等有
机溶剂。其化学组成如下:
chla: C55H72O5N4Mg
chlb: C55H70O6N4Mg
叶绿素是叶绿酸的酯。叶绿酸是双羧酸,其羧基中的
羟基分别被甲醇和叶绿醇所酯化。所以其分子式为:
看下图下一页叶绿素b以-CHO
代替-CH3图3-3 叶绿素a的结构式CH3返回4个吡咯环和4个甲烯基连成一个大环—卟啉环镁原子居卟啉环的中央1个含羰基和羧基的副环(同素环Ⅴ),羧基以酯键和甲醇结合叶绿醇则以酯键与在第Ⅳ吡珞环侧键上的丙酸结合庞大的共轭体系,起着吸收光能,传递电子,以诱导共振的方式传递能量,但不参与H的传递或氧化还原疏水尾部H+,Cu2+可取代Mg⑵胡萝卜素和叶黄素:四萜类、有α- 、β-、γ- 三种异构体。不溶于水,但能溶于有机溶剂。
①胡萝卜素:
是一不饱和的碳氢化合物,分子式为C40H56。它的两头
具有一个对称排列的紫罗兰酮环,它们中间以共轭双键
(4个异戊二烯)相联接。
②叶黄素由胡萝卜素衍生而来,分子式为C40H56O2,是
个醇类物质,它在叶绿体的结构中与脂类物质相结合。
⑶藻胆素
藻类进行光合作用的主要色素,不溶于有机溶剂,溶于
水。常与蛋白质结合为藻胆蛋白(藻红蛋白和藻蓝蛋白)。
β-胡萝卜素叶黄素图3-4 β-胡萝卜素和叶黄素结构式3、光合色素的光学特性
⑴辐射能量
光波是一种电磁波,对光合作用有效的可见光的波长是400~700nm之间。光同时又是运动着的粒子流,这些粒子称为光子,或光量子。光子携带的能量和光的波长的关系如下:E=N h c/λ
E=(6.02×1023)×(6.6262×10-34)×频率=( ) ×
阿伏伽德罗常数 普朗克常数
上式表明:光子的能量与波长成反比。
⑵吸收光谱
太阳光谱①叶绿素的吸收光谱
叶绿素吸收光的能力很强,如果把叶绿素溶液放在
光源和分光镜之间,就可以看到有些波长的光线被吸
收了。在光谱中就出现了暗带,这种光谱叫吸收光谱。
两个最强烈的吸收区,一个是波长为640~660的红
光部分,另一个是430~450的蓝紫光部分。此外,在光
谱的橙光,黄光和绿光部分只有不明显的吸收带,其
中尤以对绿光的吸收最少,所以叶绿素的溶液呈绿色。
chla和chlb的吸收光谱很相似,但略有不同。
②类胡萝卜素的的吸收光谱
最大吸收在蓝紫光部分,不吸收红光等波长的光。
叶绿素b叶绿素a图3-7 叶绿素a和叶绿素b在乙醇溶液中的吸收光谱图3-8 α-胡萝卜素和叶黄素的吸收光谱叶黄素α-胡萝卜素λ/nm返回⑶作用光谱
指在能量相同而波长不同的光下,测定其光合强
度所得的变化曲线。
作用光谱与叶绿素a的吸收光谱基本一致,说明光合作用吸
收的光一般是由叶绿素a吸收的,其它色素吸收的光都传递给叶
绿素a,然后引起光化学反应。
⑷荧光现象和磷光现象
荧光现象:叶绿素溶液在透射光下呈绿色,而在反
射光下呈红色的现象。10-8~10-9秒(寿命短)
磷光现象:叶绿素除了在照光时能辐射出荧光外,当去掉光源后,还能继续辐射出极微弱的红光(用精密仪器可测知),这个现象叫~。 10-2秒(寿命长)
这两种现象说明叶绿素能被光激发,而被光激发是将光能转变为化学能的第一步。
图3-10 色素分子吸收光子后能量转变E1E2E0第二单线态第一单线态第一三线态荧光磷
光
4、叶绿素的合成
⑴ 合成原料:谷氨酸、α- 酮戊二酸
⑵需氧气和光
⑶矿质营养 N 、 Mg ; Fe 、 Mn、 Cu、 Zn
⑷温度
⑸水分 影响蛋白质合成
谷氨酸或α- 酮戊二酸 氨基酮戊二酸(ALA)
2ALA 含吡咯环的胆色素原
4个胆色素原 尿卟啉原Ⅲ 粪吡啉原Ⅲ
粪吡啉原Ⅲ 原卟啉Ⅸ 镁原卟啉 原脱植基
叶绿素a 脱植基叶绿素a 叶绿素a 叶绿素b
厌氧条件下有氧条件下知道: 叶绿素与血红素有共同的前期合成途径。推断:动植物有共同的起源第二节 光合作用机理光合作用可分为三个阶段:
①原初反应②电子传递和光合磷酸化 光反应
③碳同化 暗反应
一、原初反应(Primary reaction):光能 电能
包括光能的吸收、传递和转换过程。
1、作用中心色素分子
指具有光学活性的特殊状态存在的少数叶绿素a分子。
光学活性:指这种色素吸收光能之后即被激发,可引起自身的 氧化还原反应(得失电子),同时将接受的光能转变成电能的性质。
特殊状态:以水结合起来的叶绿素a的二聚体,吸收高峰为
680nm,700nm。原初反应电子传递和光合磷酸化碳同化光反应暗反应2、聚光色素
除作用中心色素以外的大多数叶绿素a、全部叶绿素b、类胡萝卜素。它们都只能将吸收的光聚集起来传递给作用中心色素,它们无光学活性,只能捕捉光能,因此叫聚光色素。它们又象收音机的天线一样,因此又叫“天线色素”。
两种色素共同作用,才能将光能 电能
3、反应中心
指在类囊体中进行光合作用原初反应的最基本的色素蛋白结构。
反应中心=作用中心色素分子+原初电子供体+原初电子受体
原初电子供体:以电子直接供给作用中心色素分子的物体。
原初电子受体:直接接受作用中心色素分子传来电子的物体。
D·P·A D·P*·A D·P+·A- D+·P·A-4、光合单位
指结合在类囊体膜上能进行光合作用的最小的结构单位。
光合单位=聚光色素系统+反应中心
光合作用的原初反应是连续不断的进行的,因此,
必须有不断的最终电子供体和最终电子受体,构成电子
的“源”和“流”。高等植物的最终电子供体是水,最终电
子受体为NADP。聚光色素以诱导共振方式传递光量子,
最后传递给反应中心色素分子,这样作用中心色素被激
发而完成光能转换为电能的过程。
图3-12 光合作用原初反应的能量吸收、传递和转换反应中心H2ONADP+e
返回二、电子传递和光合磷酸化 电能 活跃的化学能 ㈠、光系统 两个光系统
量子产额:吸收一个光量子后放出的氧分子数目或固定CO2分子数目。
红降现象
以绿藻和红藻为材料,研究其不同光波的光合效率,发现当用光波大于685nm (远红光)的光照射时,虽然仍被叶绿素大量吸收,但量子产额急剧下降,这种现象被称为~。
双光增益现象(爱默生效应)
爱默生等发现,在用远红光(光波大于685nm )照射条件下,如补充红光(约650nm) ,则量子产额大增,比这两种波长的光单独照射的总和还要多。这种两种光波促进光合效率的现象,叫双光增益现象(爱默生效应)。
所以,认为光合作用包括两个光系统,后来证明确实如此。
⒈光系统Ⅰ(PSⅠ)
这是一种直径较小的颗粒,直径为11nm,存在于基质片
层和基粒片层的非垛叠区。
⒉光系统Ⅱ(PSⅡ):
这是一种直径较大的颗粒,直径为17.5nm,存在于类囊体膜的垛叠区。
Tyr p680 p680* pheo QA QB PQH2 PC
与OEC联系,水解放氧
2H2O O2 + 4H++4e-
光补充1、希尔反应(Hill reaction)
希尔发现在离体的叶绿体(实际是被膜破裂的叶绿体)悬浮液中加入适当的电子受体(如草酸铁),照光时可使水分解而释放氧气,这个反应称为希尔反应。其中的受体被称为希尔氧化剂。
水的光解和放氧:
Kok(1970)提出放氧系统的5个S状态循环模式
水氧化钟: S0,S1,S2,S3,S4表示不同氧化还原状态,每闪光一次则有不同的状态。从S0 S4每两个状态之间的转变都失去1个电子,到S4时共积累4个正电荷。 S4不稳定,从2分子水中获得4个电子又回到S0状态,同时产生O2,如此循环,每1个循环吸收4个光量子,氧化2个水分子,向PSⅡ反应中心传递4个电子并释放4个质子和1个氧分子。这种循环称水氧化钟。
电子传递和质子传递光合链:由PSⅡ、PSⅠ和Cytb6f构成
是光合作用中的电子传递系统,由两个光系统和一系列电子传递体和递氢体按照各自氧化还原电位的高低排列在光合膜上。
电子传递自发进行。图 图3-14 类囊体膜上的电子传递PSIIPSICytb6fP680+ P680*Tyr PheoP700*
P700+基质类囊体腔类囊体膜低
(H+)高
(H+)图3-15 PSII, Cytb6f复合体PSI 和ATP合酶复合体中的电子和质子传递PSIIPSI ㈡、光合磷酸化( Photophosphorylation ):叶绿体在光下把无机磷和ADP转化为ATP,形成高能磷酸键的过程,称光合磷酸化。
1、光合磷酸化形式
(1)非环式光合磷酸化
OEC水解后,把H+释放到类囊体腔,把电子释放到PSⅡ内,
电子在光合电子传递链中传递时,伴随着类囊体外侧的H+转移
到腔内,由此形成了跨膜的H+浓度差,引起ATP的形成,同时
把电子传递到PSⅠ去,进一步提高了能位,而使H+还原NADP+
为NADPH,此外还放出O2 ,在这个过程中,电子传递是一个开
放的通路,在此通路中伴随着ATP的形成,故称~。
2ADP+2Pi+2NADPH+2H2O 2ATP+2NADPH+O2
如图 图 图3-14 类囊体膜上的电子传递PSIIPSICytb6fP680+ P680*Tyr PheoP700*
P700+基质类囊体腔类囊体膜低
(H+)高
(H+)图3-15 PSII, Cytb6f复合体PSI 和ATP合酶复合体中的电子和质子传递PSIIPSI(2)环式光合磷酸化
PSⅠ产生的电子经过一些电子传递体后,伴随形成腔内外H+浓度差,只引起ATP的形成,不放出O2,也无NADP+ 的还原反应,在这个过程中,电子经过一系列传递后降低了能位,最后经过PC重新回到原来的起点,也就是电子的传递是一个闭合的回路,故称环式光合磷酸化。图 图3-14 类囊体膜上的电子传递PSIIPSICytb6fP680+ P680*Tyr PheoP700*
P700+基质类囊体腔类囊体膜低
(H+)高
(H+)图3-15 PSII, Cytb6f复合体PSI 和ATP合酶复合体中的电子和质子传递PSIIPSI(3)假环式光合磷酸化
以O2代替NADP +作为末端电子受体,其发生磷酸化的部位与非环式相同,在NADP供应量较低时,有利于此过程的进行。
2、ATP合酶
ATP合酶位于基质片层和基粒片层的非垛叠区,这个复合物的功能是把ADP和Pi合成为ATP,故名ATP合酶。它将ATP的合成与电子传递和H+跨膜转运偶联起来,故又称为偶联因子。
ATP合酶复合体有亲水性头部(CF1)和疏水性柄部(CF0)组成。类囊体腔中的H+可进入CF0 移动到CF1,被位于顶部的ATP合酶催化合成ATP。下一页3、光合磷酸化机理化学渗透假说(Chemiosmotic hypothesi)
在光下,由于光合色素吸收光量子,传至反应中心色素而引起光化学反应及电子传递,PQ在接受2电子的同时,又从膜外间质中获得2质子而成还原态(PQH2), PQH2有将电子传给Cytb和Fe-S,由于Cytb和Fe-S只接受电子而不接受质子,于是H+释放在膜内(类囊体空间),同时水在膜内侧也释放H+,因而形成跨膜的H+梯度(浓度差),膜内H+浓度高而膜外则降低,膜内电位正、膜外较负(电位差)。二者合称为电化学梯度,也是光合磷酸化的动力(质子动力)。 H+有沿着电化学梯度返回膜外的趋势,当通过ATP酶复合物时即做功,使ADP+Pi ATP,这就是光合磷酸化。基质类囊体腔类囊体膜低
(H+)高
(H+)图3-15 PSII, Cytb6f复合体PSI 和ATP合酶复合体中的电子和质子传递PSIIPSI 经上述变化后
光能 电能 活跃的化学能(ATP,NADPH)
ATP和NADPH只能暂时存在而不能积累,还要
向下传递,NADPH的H又能进一步还原CO2固定形成
的中间产物。这样就把光反应和暗反应联系起来了。
叶绿体有了ATP和NADPH,就可在暗反应中同
化CO2 。所以, ATP和NADPH 又叫“同化能力” 、
“同化力” 、“同化功”。
到此,光合作用的光反应进行完毕,下面我们讲
暗反应(碳同化)。
三、碳同化(CO2 assimilation)活跃的化学能 稳定的化学能
在叶绿体间质进行
① C3 途径(卡尔文循环)
② C4 途径
③景天酸代谢(CAM)
固定转移CO2,能形成淀粉等产物
㈠ C3 途径(卡尔文循环)
卡尔文等利用放射性同位素和纸层析等方法,提出二氧化碳同化的循环途径,故称卡尔文循环或光合环。由于这个循环中的CO2 受体是一种戊糖(核酮糖二磷酸),故又称为还原戊糖磷酸途径。这个途径的CO2固定最初产物是一种三碳化合物,故又称C3途径。
C3途径大致可分为三个阶段:
①羧化阶段,②还原阶段,③更新阶段
固定转移CO2,能形成淀粉等产物⒈羧化阶段( Carboxylation phase)
RuBP +CO2 2PGA
⒉还原阶段 (Retuction phase)
PGA D-PGA PGAld
从PGA 到PGAld 过程中,由光合作用生成的ATP与 NADPH均被利用掉。CO2一旦还原到PGAld,光合作用的储能过程便完成。PGAld等三碳糖可进一步在叶绿体内合成淀粉,也可透出叶绿体,在细胞质中合成蔗糖。
CO2受体初产物⒊更新阶段(Regeneration phase)
RuBP图3-17 卡尔文循环①是羧化阶段 ②,③ 是还原阶段,其余反应是更新阶段①②③PGA3CO26ATP6ADP6NADPH6NADP+6PiDPGAPGAldDHAP 3CO2+3H2O+3RuBP+9ATP+6NADPH
PGAld+6NADP++9ADP+9Pi
⒋ C3 途径的调节
⑴酶活性的调节
光 ,H+, pH , Mg
⑵质量作用的调节
代谢物浓度影响反应的方向和速度。
⑶转运作用的调节
磷酸转运器
㈡ C4途径(四碳二羧酸途径) 1. C3与C4植物 据植物同化CO2 途径分为: C3植物:把仅通过C3途径就可以完成CO2固定作用的植物称为C3植物。多为温带和寒温带植物,如小麦、大豆、棉花、油菜等。 C4植物:需C3 和C4两条途径才能完成CO2的同化,这样的植物称为C4植物。多为热带和亚热带植物,如玉米、高粱、甘蔗、苋菜等。 CAM植物:通过景天酸代谢途径完成CO2的同化,这样的植物称为CAM植物。一般生长在干旱条件下,气孔晚上开,白天关,节水。 C3与C4植物的解剖学差异 ① C4植物中柱鞘有叶绿体, ② C4植物有花环状结构, ③ 中柱鞘细胞与叶肉细胞有大量的胞间连丝联系。 ④叶肉细胞排列情况不同, C3植物排列松散, C4 植物排列紧密。C3植物C4植物PEP+HCO3- OAA+Pi 发生在细胞质中(如图)
发生部位、酶、受体、产物与C3 均不同。 PEPC 四碳二羧酸CO2受体2、CO2的固定C4途径C3途径叶肉细胞维管束细胞图3-18 C4植物的C4途径和C3途径发生部位草酰乙酸 CO2PEPOAA 的进一步转化因植物种类而异(如图)
NADP苹果酸酶型
NAD苹果酸酶型
PEP羧激酶型叶肉细胞维管束鞘细胞苹果酸天冬氨酸天冬氨酸NADP苹果酸酶型NAD苹果酸酶型PEP羧激酶型 图3-19 C4植物的3种类型-
脱羧部位是叶绿体
NADP苹果酸酶
返回的是丙酮酸脱羧部位是线粒体
NAD苹果酸酶
返回的是丙氨酸脱羧部位是细胞质
PEP羧激酶
返回的是丙酮酸和丙氨酸㈢ CAM途径(景天科酸代谢途径)仙人掌,落地生根,菠萝,兰花等
气孔白天关闭,夜间气孔开放,固定二氧化碳。
夜间气孔开放 PEP+CO2 OAA+Pi
OAA 苹果酸 积累在液泡
白天气孔关闭 苹果酸 丙酮酸+CO2
PEP主要来自于呼吸作用的PPP途径和EMP途径。
这样,呼吸作用和光合作用相互联系起来。
调节
短期调节: 夜晚型和白天型
长期调节:兼性或诱导性CAM植物PEPC苹果酸脱氢酶苹果酸酶NADPNADPH发生在细胞质发生在叶绿体液泡叶绿体细胞质第三节 光呼吸 (Photorespiration)光呼吸
植物的绿色细胞在光下吸收O2,释放CO2的过程。
一、光呼吸的生化历程
光呼吸是一个氧化过程,被氧化的底物是乙醇酸。
㈠ 光呼吸底物乙醇酸的生物合成
RuBPC具有双重活性
RuBP + O2 2-磷酸乙醇酸 + 3-磷酸甘油酸
2-磷酸乙醇酸 乙醇酸 + 磷酸
以上是在叶绿体中进行的
㈡ 乙醇酸的氧化及CO2的产生
乙醇酸在叶绿体形成后,就转入另一个细胞器-过氧化物酶体。
C3植物叶肉细胞的过氧化物酶体较多, 而C4植物的过氧化物酶体较少,大多数在维管束鞘细胞的薄壁细胞内,一般位于叶绿体附近。 乙醇酸 + O2 乙醛酸 + H2O2
乙醛酸 + 谷氨酸 甘氨酸+α-酮戊二酸
甘氨酸转入线粒体,脱羧产生CO2
2甘氨酸 丝氨酸 + CO2 + NH3 + NADH2
丝氨酸回到过氧化物酶体,经转氨作用生成羟基丙酮酸
丝氨酸 + α-酮戊二酸 羟基丙酮酸 + 谷氨酸
羟基丙酮酸 + NADH2 甘油酸 + NAD+
甘油酸进入叶绿体,经磷酸化生成3-磷酸甘油酸,进入C3途
径,为进一步形成RuBP的光呼吸底物乙醇酸创造条件。
由于乙醇酸(底物)是C2化合物,氧化产物乙醛酸、甘氨酸
都是C2化合物,故也称光呼吸为二碳光呼吸氧化环或称C2环。 图3-26 光呼吸代谢途径线粒体 过氧化物酶体叶绿体磷酸乙醇酸NH3+CO2甘氨酸丝氨酸ATPADPNAD+NADH甘油酸二、光呼吸的特点⒈ 光呼吸随反应环境中氧浓度的增加而加速。
⒉ 光呼吸与光合作用关系密切。
⑴光呼吸底物乙醇酸来源于光合作用中的RuBP
⑵光合生成的有机物的1/3以上要消耗在光呼吸底物上
⑶光呼吸消耗能量
⒊光呼吸是在叶绿体、过氧化体、线粒体3 种细胞器联系作用下进行的。
⒋光呼吸的吸O2和排CO2不在同一个细胞器里,吸收O2是在叶绿体、过氧化体中,放出CO2在线粒体中。
三、光呼吸的生理意义
1. 防止强光对光合机构的破坏 光呼吸是一个保护性的生理过程,在高光强、低CO2 时会形成过量的同化力,由光激发的高能电子会传递给O2,形成超氧阴离子自由基会对光合膜、光合器官有伤害作用,而光呼吸可消耗同化力和高能电子,降低超氧阴离子自由基的形成,从而保护叶绿体,免除或减少强光对光合机构的破坏。
2. 维持C3循环的运转 在干旱和高辐射期间,气孔关闭,CO2不能进入,会导至光抑制,此时光呼吸释放CO2 ,能被C3途径再利用,避免产生光抑制。
3. 回收碳素 通过二碳氧化环可回收3/4的碳(2个乙醇酸转化1个PGA,释放1个CO2 ) 4. 对蛋白质合成提供了甘氨酸和丝氨酸。 5. 能消除乙醇酸的毒害作用。光呼吸可以转化乙醇酸。 6.可以为硝酸盐的还原提供还原型NAD四、比较C3植物和C4植物的光合特征 ㈠结构
㈡生理
⒈ 酶
⒉ CO2的补偿点
⒊ 光呼吸
⒋ 利用组织中CO2的情况
⒌ 热限
⒍ 所需能量
⒈酶对CO2的亲和力
PEPC 对CO2的Km = 7umol
RuBPC 对CO2的Km = 450umol
所以, PEPC对CO2的亲和力大。
⒉ CO2的补偿点
定义:指在照光情况下,植物光合作用所吸收的CO2量与呼吸作用所释放的CO2量达到动态平衡时外界环境中CO2浓度。
C3 植物CO2 补偿点高, C4 植物CO2 补偿点低。
⒊光呼吸
C3 植物是高光呼吸植物, C4 植物是低光呼吸植物
⑴ PEPC 对CO2亲和力要高于RuBPC
⑵ C4 植物RuBPC位于维管束鞘细胞,O2浓度较小;
⑶ 四碳二羧酸从叶肉细胞进入鞘细胞,起到“CO2泵”的作用,增加鞘细胞CO2浓度,增加RuBP 的羧化反应,减少了加氧反应,
⑷ C4植物PEP羧化酶位于叶肉细胞,对光呼吸释放的CO2 有捕捉作用
因此C4植物光呼吸小。
⒋利用组织中CO2的情况
C4植物①能利用细胞间隙中较低浓度的CO2
②光呼吸放出的CO2不易漏出。
⒌热限
C4植物热限比C3植物的热限高。
⒍所需能量
C4植物需额外的能量用于PEP的再生。
第四节 光合作用产物(photosynthetic product)主要是蔗糖和淀粉
一、淀粉的合成
(1)场所:叶绿体
(2)原料:来自于C3途径的磷酸丙糖
DHAP+PGAld FBP F6P G6P G1P
G1P+ATP ADPG
( G)n
二、蔗糖的合成
(1)场所:细胞质
(2)原料:来自于C3途径的磷酸丙糖
(3)磷酸运送器(Phosphate translocator)
磷酸运送器(Phosphate translocator)
是在叶绿体和胞质溶胶之间转运磷酸的专一载体。每从叶绿体运出一个磷酸酯就有一个正磷酸离子从胞质溶胶运送回来。
磷酸丙糖 + Pi 磷酸丙糖 + Pi
(叶绿体) (细胞质) (细胞质) (叶绿体)
DHAP+PGAld FBP F6P G6P G1P
G1P+UTP UDPG
F6P
三、蔗糖和淀粉合成的调节
由磷酸和磷酸丙糖的相对浓度控制。
Pi, F6P促进蔗糖合成。磷酸运送器蔗糖-6-磷酸蔗糖合酶蔗糖第五节 影响光合作用的因素一、光合作用的指标
1、光合强度(Intensity of photosynthesis)
也称光合速率 (Photosynthetic rate)
单位时间、单位叶面积吸收CO2的量或放出O2的量。
2、净光合强度(表观光合强度)
实际所测得的光合速率
半叶法、红外CO2分析仪法、氧电极法
3、总光合速率(Gross Photosynthetic rate )
也称真光合速率(Ture Photosynthetic rate )
表观光合速率加上光、暗呼吸速率
二、影响光合作用的因素1、光照 : 光强、光质
2、CO2浓度
3、矿质营养
4、水分
5、温度:三基点温度
6、光合作用的日变化1、光照 : 光强、光质
⑴光补偿点(Light compensation point)
同一叶子在同一时间内,光合过程中吸收的CO2和呼吸过程中放出的CO2等量时的光照强度。
⑵光饱和现象(Light saturation) /光饱和点(Light saturation point)
当达到某一光强时,光合速率就不再随光强的增强而增加,这种现象称为光饱和现象。
刚刚出现光饱和现象时的光照强度称为光饱和点。
⑶光抑制(Photoinhibition)
光能超过光合系统所能利用的时,光合功能下降的现象叫~。
⑷ 光质:红光最有效2、CO2浓度
⑴ CO2饱和点(CO2 Saturation point)
在一定范围内,植物净光合速率随CO2浓度升高而增加,但到达一定程度时,再增加CO2浓度,净光合速率不再增加,这时CO2浓度叫CO2饱和点。
⑵ CO2补偿点(CO2 Compensation point)
降低环境温度可以降低光补偿点。(??? ) CO2的补偿点随光照强度的变化而变化。(??? )CO2饱和度随光照强度的增加而升高.(??? ) 3、矿质营养
直接、间接影响光合作用
调节气孔开度,影响碳水化合物的转换及运输,光合链成分,叶绿素组分,酶的活化剂,加大光合面积等。4、水分
⑴ 缺水引起气孔开度小或关闭,限制了CO2的吸收----气孔阻力
⑵ 缺水时,淀粉水解加强,糖类堆积,光合产物输出缓慢,抑制光合作用的进行,而促进了呼吸作用。
(3)缺水减少光合面积。
(4)缺水会降低CO2在细胞间隙内的传递速度---叶肉阻力。
5、温度:三基点温度
C4植物的最适温度高于C3植物
C4植物的热限高于C3植物
6、光合作用的日变化
⑴ 温暖的日子,水分充足,太阳光是主要矛盾,光合速率与太阳辐射进程相符合。
⑵ 如白天云量变化不定,光合速率呈不规则曲线。
⑶晴天无云,但光强强烈时,“午休”现象。
光合“午休”现象(midday depression)
指植物的光合速率在中午前后下降的现象。引起“午休”的主要原因是大气干旱和土壤干旱及强光、高温、低CO2浓度等。
第六节 植物对光能的利用一、光能利用率
指植物光合作用所积累的有机物所含的能量,占照射在单位地面上的日光能的比率。一般为5%。
1、量子需要量:每同化1分子CO2或释放 1分子O2所需的光量子数。
2、量子产额或量子效率:每吸收1个光量子所能同化的CO2或释放 的O2的分子数。
3、生物学产量:光合产量-光合产物消耗。
4、光合产量:光合面积×光合强度×光合时间
5、经济产量:生物学产量×经济系数
6、经济系数:指收获物中经济价值较高的部位的产量占作物总
量的比值。
光能利用率低的原因: 不能吸收的波长 60%,
反射和透光 8%
热能散失 8%
代谢 19%
转化 5%二、光合产物的分配规律
由“代谢源”到“代谢库”
代谢源:指制造和输出同化物的部位,如成熟的叶片。
代谢库:指输出和利用同化物的部位,如果实、种子。
⒈就近供应
⒉优先供应生长中心
⒊同侧供应
提高植物光能利用率的途径
经济产量=(光合面积×光合速率×光合时间-消耗)×经济系数
延长光照时间
提高复种指数(全年内作物收获面积与耕地面积之比):套种或间作。
延长生长期:如育苗移栽、套种、适时早播、防止早衰。
补充人工光照
增加光合面积
合理密植
改善株型:杆矮、叶直而小、叶片厚、分蘖密集。
加强光合效率
提高CO2浓度:通风、施CO2(干冰)、施有机肥、碳铵
抑制光呼吸:光呼吸抑制剂,如α-羟基磺酸可抑制乙醇酸氧化酶活性。
提高经济系数(经济产量与生物产量之比)测试1、在光照强,温度高,空气湿度低的情况下, C3 和C4植物哪一个光合效率高?为什么?(光饱和点,热限,CO2泵, CO2补偿点, CO2亲和力,光呼吸,利用组织CO2情况)
2、在光照弱,温度低,空气湿度高的情况下, C3 和C4植物哪一个光合效率高?为什么?
3、在碳同化中受光调节的酶有哪几种?
4、如何证明光合作用中释放的氧气来自水。
5、如何证明光合电子传递有两个光系统参与?
6、在缺乏CO2的情况下,对绿色叶片照光能观察到荧光,然后在供给CO2的情况下,荧光立即被淬灭,试解释其原因。
补充:叶绿素的理化性质⑴ 荧光现象和磷光现象
⑵ 皂化现象
⑶H,Cu代镁现象
+2KOH+ CH3OH+C20H39OH补充:叶绿素的层析
胡萝卜素叶黄素叶绿素a叶绿素b
光合作用的意义和研究历史
叶绿体和光合色素
光合作用机理
光呼吸
影响光合速率的外界因素
光合作用与农业生产
概述
一、自养植物和异养植物
1、异养植物(Heterophyte)
2、自养植物(Autophyte)
二、碳素同化作用(Carbon assimilation)
1、光合作用(Photosynthesis)
光
CO2 +H2O (CH2O ) + O2
叶绿体什么是光合作用? 绿色植物在光下,把二氧化碳和水转化为糖,并释放出氧气的过程。
厂房叶绿体动力光能原料二氧化碳和水产物有机物和氧 2、细菌光合作用 (Bacterial photosynthesis)
光、叶绿素
CO2 + H2S CH2O + H2O+S
3、化能合成作用(Chemosynthesis)
化能合成细菌
三、光合作用的重要性
1、有机物质的重要来源
2、把光能转化成化学能
3、大气中氧气的重要来源
第一节 叶绿体和叶绿体色素I一、叶绿体(Chloroplast)
㈠叶绿体的结构:
椭圆形,一般直径为3~6um,厚为2~3um。每平方毫米的蓖麻叶就含3~5百万个叶绿体。
1、叶绿体膜—选择性屏障,控制物质进出。
2 、基质—CO2的固定,淀粉的合成和储藏(含酶类)
3 、基粒—光能-化学能(光合色素)
4 、嗜锇滴—基质中与锇酸容易结合的颗粒(醌类)
5 、类囊体—光合作用能量转换(又称光合膜)
基粒类囊体 (grana thylakoid)
基质类囊体 (stroma thylakoid)下一页外膜内膜基质基粒㈡叶绿体的成分
1、 水分(75%)
2 、 蛋白质(30~45%)—催化剂
3 、脂类(20~40%)—膜成分
4 、色素(8%)—与蛋白质结合,电子传递
5 、无机盐(10%)
6 、储藏物质(如淀粉等,10~20%)
7 、NAD+、NADP+、醌(如质体醌),起传递氢原子或电子的作用。二、光合色素
1、分类
叶绿素:类胡萝卜素=3:1 所以叶片一般呈绿色
叶绿素a:叶绿素b=3:1
叶黄素:胡萝卜素=2:1
解释:秋后或衰老的叶片多呈黄色,秋后枫树叶子呈红色2、光合色素化学结构与性质
⑴叶绿素(chlorophyll)
叶绿素不溶于水,但能溶于酒精、丙酮和石油醚等有
机溶剂。其化学组成如下:
chla: C55H72O5N4Mg
chlb: C55H70O6N4Mg
叶绿素是叶绿酸的酯。叶绿酸是双羧酸,其羧基中的
羟基分别被甲醇和叶绿醇所酯化。所以其分子式为:
看下图下一页叶绿素b以-CHO
代替-CH3图3-3 叶绿素a的结构式CH3返回4个吡咯环和4个甲烯基连成一个大环—卟啉环镁原子居卟啉环的中央1个含羰基和羧基的副环(同素环Ⅴ),羧基以酯键和甲醇结合叶绿醇则以酯键与在第Ⅳ吡珞环侧键上的丙酸结合庞大的共轭体系,起着吸收光能,传递电子,以诱导共振的方式传递能量,但不参与H的传递或氧化还原疏水尾部H+,Cu2+可取代Mg⑵胡萝卜素和叶黄素:四萜类、有α- 、β-、γ- 三种异构体。不溶于水,但能溶于有机溶剂。
①胡萝卜素:
是一不饱和的碳氢化合物,分子式为C40H56。它的两头
具有一个对称排列的紫罗兰酮环,它们中间以共轭双键
(4个异戊二烯)相联接。
②叶黄素由胡萝卜素衍生而来,分子式为C40H56O2,是
个醇类物质,它在叶绿体的结构中与脂类物质相结合。
⑶藻胆素
藻类进行光合作用的主要色素,不溶于有机溶剂,溶于
水。常与蛋白质结合为藻胆蛋白(藻红蛋白和藻蓝蛋白)。
β-胡萝卜素叶黄素图3-4 β-胡萝卜素和叶黄素结构式3、光合色素的光学特性
⑴辐射能量
光波是一种电磁波,对光合作用有效的可见光的波长是400~700nm之间。光同时又是运动着的粒子流,这些粒子称为光子,或光量子。光子携带的能量和光的波长的关系如下:E=N h c/λ
E=(6.02×1023)×(6.6262×10-34)×频率=( ) ×
阿伏伽德罗常数 普朗克常数
上式表明:光子的能量与波长成反比。
⑵吸收光谱
太阳光谱①叶绿素的吸收光谱
叶绿素吸收光的能力很强,如果把叶绿素溶液放在
光源和分光镜之间,就可以看到有些波长的光线被吸
收了。在光谱中就出现了暗带,这种光谱叫吸收光谱。
两个最强烈的吸收区,一个是波长为640~660的红
光部分,另一个是430~450的蓝紫光部分。此外,在光
谱的橙光,黄光和绿光部分只有不明显的吸收带,其
中尤以对绿光的吸收最少,所以叶绿素的溶液呈绿色。
chla和chlb的吸收光谱很相似,但略有不同。
②类胡萝卜素的的吸收光谱
最大吸收在蓝紫光部分,不吸收红光等波长的光。
叶绿素b叶绿素a图3-7 叶绿素a和叶绿素b在乙醇溶液中的吸收光谱图3-8 α-胡萝卜素和叶黄素的吸收光谱叶黄素α-胡萝卜素λ/nm返回⑶作用光谱
指在能量相同而波长不同的光下,测定其光合强
度所得的变化曲线。
作用光谱与叶绿素a的吸收光谱基本一致,说明光合作用吸
收的光一般是由叶绿素a吸收的,其它色素吸收的光都传递给叶
绿素a,然后引起光化学反应。
⑷荧光现象和磷光现象
荧光现象:叶绿素溶液在透射光下呈绿色,而在反
射光下呈红色的现象。10-8~10-9秒(寿命短)
磷光现象:叶绿素除了在照光时能辐射出荧光外,当去掉光源后,还能继续辐射出极微弱的红光(用精密仪器可测知),这个现象叫~。 10-2秒(寿命长)
这两种现象说明叶绿素能被光激发,而被光激发是将光能转变为化学能的第一步。
图3-10 色素分子吸收光子后能量转变E1E2E0第二单线态第一单线态第一三线态荧光磷
光
4、叶绿素的合成
⑴ 合成原料:谷氨酸、α- 酮戊二酸
⑵需氧气和光
⑶矿质营养 N 、 Mg ; Fe 、 Mn、 Cu、 Zn
⑷温度
⑸水分 影响蛋白质合成
谷氨酸或α- 酮戊二酸 氨基酮戊二酸(ALA)
2ALA 含吡咯环的胆色素原
4个胆色素原 尿卟啉原Ⅲ 粪吡啉原Ⅲ
粪吡啉原Ⅲ 原卟啉Ⅸ 镁原卟啉 原脱植基
叶绿素a 脱植基叶绿素a 叶绿素a 叶绿素b
厌氧条件下有氧条件下知道: 叶绿素与血红素有共同的前期合成途径。推断:动植物有共同的起源第二节 光合作用机理光合作用可分为三个阶段:
①原初反应②电子传递和光合磷酸化 光反应
③碳同化 暗反应
一、原初反应(Primary reaction):光能 电能
包括光能的吸收、传递和转换过程。
1、作用中心色素分子
指具有光学活性的特殊状态存在的少数叶绿素a分子。
光学活性:指这种色素吸收光能之后即被激发,可引起自身的 氧化还原反应(得失电子),同时将接受的光能转变成电能的性质。
特殊状态:以水结合起来的叶绿素a的二聚体,吸收高峰为
680nm,700nm。原初反应电子传递和光合磷酸化碳同化光反应暗反应2、聚光色素
除作用中心色素以外的大多数叶绿素a、全部叶绿素b、类胡萝卜素。它们都只能将吸收的光聚集起来传递给作用中心色素,它们无光学活性,只能捕捉光能,因此叫聚光色素。它们又象收音机的天线一样,因此又叫“天线色素”。
两种色素共同作用,才能将光能 电能
3、反应中心
指在类囊体中进行光合作用原初反应的最基本的色素蛋白结构。
反应中心=作用中心色素分子+原初电子供体+原初电子受体
原初电子供体:以电子直接供给作用中心色素分子的物体。
原初电子受体:直接接受作用中心色素分子传来电子的物体。
D·P·A D·P*·A D·P+·A- D+·P·A-4、光合单位
指结合在类囊体膜上能进行光合作用的最小的结构单位。
光合单位=聚光色素系统+反应中心
光合作用的原初反应是连续不断的进行的,因此,
必须有不断的最终电子供体和最终电子受体,构成电子
的“源”和“流”。高等植物的最终电子供体是水,最终电
子受体为NADP。聚光色素以诱导共振方式传递光量子,
最后传递给反应中心色素分子,这样作用中心色素被激
发而完成光能转换为电能的过程。
图3-12 光合作用原初反应的能量吸收、传递和转换反应中心H2ONADP+e
返回二、电子传递和光合磷酸化 电能 活跃的化学能 ㈠、光系统 两个光系统
量子产额:吸收一个光量子后放出的氧分子数目或固定CO2分子数目。
红降现象
以绿藻和红藻为材料,研究其不同光波的光合效率,发现当用光波大于685nm (远红光)的光照射时,虽然仍被叶绿素大量吸收,但量子产额急剧下降,这种现象被称为~。
双光增益现象(爱默生效应)
爱默生等发现,在用远红光(光波大于685nm )照射条件下,如补充红光(约650nm) ,则量子产额大增,比这两种波长的光单独照射的总和还要多。这种两种光波促进光合效率的现象,叫双光增益现象(爱默生效应)。
所以,认为光合作用包括两个光系统,后来证明确实如此。
⒈光系统Ⅰ(PSⅠ)
这是一种直径较小的颗粒,直径为11nm,存在于基质片
层和基粒片层的非垛叠区。
⒉光系统Ⅱ(PSⅡ):
这是一种直径较大的颗粒,直径为17.5nm,存在于类囊体膜的垛叠区。
Tyr p680 p680* pheo QA QB PQH2 PC
与OEC联系,水解放氧
2H2O O2 + 4H++4e-
光补充1、希尔反应(Hill reaction)
希尔发现在离体的叶绿体(实际是被膜破裂的叶绿体)悬浮液中加入适当的电子受体(如草酸铁),照光时可使水分解而释放氧气,这个反应称为希尔反应。其中的受体被称为希尔氧化剂。
水的光解和放氧:
Kok(1970)提出放氧系统的5个S状态循环模式
水氧化钟: S0,S1,S2,S3,S4表示不同氧化还原状态,每闪光一次则有不同的状态。从S0 S4每两个状态之间的转变都失去1个电子,到S4时共积累4个正电荷。 S4不稳定,从2分子水中获得4个电子又回到S0状态,同时产生O2,如此循环,每1个循环吸收4个光量子,氧化2个水分子,向PSⅡ反应中心传递4个电子并释放4个质子和1个氧分子。这种循环称水氧化钟。
电子传递和质子传递光合链:由PSⅡ、PSⅠ和Cytb6f构成
是光合作用中的电子传递系统,由两个光系统和一系列电子传递体和递氢体按照各自氧化还原电位的高低排列在光合膜上。
电子传递自发进行。图 图3-14 类囊体膜上的电子传递PSIIPSICytb6fP680+ P680*Tyr PheoP700*
P700+基质类囊体腔类囊体膜低
(H+)高
(H+)图3-15 PSII, Cytb6f复合体PSI 和ATP合酶复合体中的电子和质子传递PSIIPSI ㈡、光合磷酸化( Photophosphorylation ):叶绿体在光下把无机磷和ADP转化为ATP,形成高能磷酸键的过程,称光合磷酸化。
1、光合磷酸化形式
(1)非环式光合磷酸化
OEC水解后,把H+释放到类囊体腔,把电子释放到PSⅡ内,
电子在光合电子传递链中传递时,伴随着类囊体外侧的H+转移
到腔内,由此形成了跨膜的H+浓度差,引起ATP的形成,同时
把电子传递到PSⅠ去,进一步提高了能位,而使H+还原NADP+
为NADPH,此外还放出O2 ,在这个过程中,电子传递是一个开
放的通路,在此通路中伴随着ATP的形成,故称~。
2ADP+2Pi+2NADPH+2H2O 2ATP+2NADPH+O2
如图 图 图3-14 类囊体膜上的电子传递PSIIPSICytb6fP680+ P680*Tyr PheoP700*
P700+基质类囊体腔类囊体膜低
(H+)高
(H+)图3-15 PSII, Cytb6f复合体PSI 和ATP合酶复合体中的电子和质子传递PSIIPSI(2)环式光合磷酸化
PSⅠ产生的电子经过一些电子传递体后,伴随形成腔内外H+浓度差,只引起ATP的形成,不放出O2,也无NADP+ 的还原反应,在这个过程中,电子经过一系列传递后降低了能位,最后经过PC重新回到原来的起点,也就是电子的传递是一个闭合的回路,故称环式光合磷酸化。图 图3-14 类囊体膜上的电子传递PSIIPSICytb6fP680+ P680*Tyr PheoP700*
P700+基质类囊体腔类囊体膜低
(H+)高
(H+)图3-15 PSII, Cytb6f复合体PSI 和ATP合酶复合体中的电子和质子传递PSIIPSI(3)假环式光合磷酸化
以O2代替NADP +作为末端电子受体,其发生磷酸化的部位与非环式相同,在NADP供应量较低时,有利于此过程的进行。
2、ATP合酶
ATP合酶位于基质片层和基粒片层的非垛叠区,这个复合物的功能是把ADP和Pi合成为ATP,故名ATP合酶。它将ATP的合成与电子传递和H+跨膜转运偶联起来,故又称为偶联因子。
ATP合酶复合体有亲水性头部(CF1)和疏水性柄部(CF0)组成。类囊体腔中的H+可进入CF0 移动到CF1,被位于顶部的ATP合酶催化合成ATP。下一页3、光合磷酸化机理化学渗透假说(Chemiosmotic hypothesi)
在光下,由于光合色素吸收光量子,传至反应中心色素而引起光化学反应及电子传递,PQ在接受2电子的同时,又从膜外间质中获得2质子而成还原态(PQH2), PQH2有将电子传给Cytb和Fe-S,由于Cytb和Fe-S只接受电子而不接受质子,于是H+释放在膜内(类囊体空间),同时水在膜内侧也释放H+,因而形成跨膜的H+梯度(浓度差),膜内H+浓度高而膜外则降低,膜内电位正、膜外较负(电位差)。二者合称为电化学梯度,也是光合磷酸化的动力(质子动力)。 H+有沿着电化学梯度返回膜外的趋势,当通过ATP酶复合物时即做功,使ADP+Pi ATP,这就是光合磷酸化。基质类囊体腔类囊体膜低
(H+)高
(H+)图3-15 PSII, Cytb6f复合体PSI 和ATP合酶复合体中的电子和质子传递PSIIPSI 经上述变化后
光能 电能 活跃的化学能(ATP,NADPH)
ATP和NADPH只能暂时存在而不能积累,还要
向下传递,NADPH的H又能进一步还原CO2固定形成
的中间产物。这样就把光反应和暗反应联系起来了。
叶绿体有了ATP和NADPH,就可在暗反应中同
化CO2 。所以, ATP和NADPH 又叫“同化能力” 、
“同化力” 、“同化功”。
到此,光合作用的光反应进行完毕,下面我们讲
暗反应(碳同化)。
三、碳同化(CO2 assimilation)活跃的化学能 稳定的化学能
在叶绿体间质进行
① C3 途径(卡尔文循环)
② C4 途径
③景天酸代谢(CAM)
固定转移CO2,能形成淀粉等产物
㈠ C3 途径(卡尔文循环)
卡尔文等利用放射性同位素和纸层析等方法,提出二氧化碳同化的循环途径,故称卡尔文循环或光合环。由于这个循环中的CO2 受体是一种戊糖(核酮糖二磷酸),故又称为还原戊糖磷酸途径。这个途径的CO2固定最初产物是一种三碳化合物,故又称C3途径。
C3途径大致可分为三个阶段:
①羧化阶段,②还原阶段,③更新阶段
固定转移CO2,能形成淀粉等产物⒈羧化阶段( Carboxylation phase)
RuBP +CO2 2PGA
⒉还原阶段 (Retuction phase)
PGA D-PGA PGAld
从PGA 到PGAld 过程中,由光合作用生成的ATP与 NADPH均被利用掉。CO2一旦还原到PGAld,光合作用的储能过程便完成。PGAld等三碳糖可进一步在叶绿体内合成淀粉,也可透出叶绿体,在细胞质中合成蔗糖。
CO2受体初产物⒊更新阶段(Regeneration phase)
RuBP图3-17 卡尔文循环①是羧化阶段 ②,③ 是还原阶段,其余反应是更新阶段①②③PGA3CO26ATP6ADP6NADPH6NADP+6PiDPGAPGAldDHAP 3CO2+3H2O+3RuBP+9ATP+6NADPH
PGAld+6NADP++9ADP+9Pi
⒋ C3 途径的调节
⑴酶活性的调节
光 ,H+, pH , Mg
⑵质量作用的调节
代谢物浓度影响反应的方向和速度。
⑶转运作用的调节
磷酸转运器
㈡ C4途径(四碳二羧酸途径) 1. C3与C4植物 据植物同化CO2 途径分为: C3植物:把仅通过C3途径就可以完成CO2固定作用的植物称为C3植物。多为温带和寒温带植物,如小麦、大豆、棉花、油菜等。 C4植物:需C3 和C4两条途径才能完成CO2的同化,这样的植物称为C4植物。多为热带和亚热带植物,如玉米、高粱、甘蔗、苋菜等。 CAM植物:通过景天酸代谢途径完成CO2的同化,这样的植物称为CAM植物。一般生长在干旱条件下,气孔晚上开,白天关,节水。 C3与C4植物的解剖学差异 ① C4植物中柱鞘有叶绿体, ② C4植物有花环状结构, ③ 中柱鞘细胞与叶肉细胞有大量的胞间连丝联系。 ④叶肉细胞排列情况不同, C3植物排列松散, C4 植物排列紧密。C3植物C4植物PEP+HCO3- OAA+Pi 发生在细胞质中(如图)
发生部位、酶、受体、产物与C3 均不同。 PEPC 四碳二羧酸CO2受体2、CO2的固定C4途径C3途径叶肉细胞维管束细胞图3-18 C4植物的C4途径和C3途径发生部位草酰乙酸 CO2PEPOAA 的进一步转化因植物种类而异(如图)
NADP苹果酸酶型
NAD苹果酸酶型
PEP羧激酶型叶肉细胞维管束鞘细胞苹果酸天冬氨酸天冬氨酸NADP苹果酸酶型NAD苹果酸酶型PEP羧激酶型 图3-19 C4植物的3种类型-
脱羧部位是叶绿体
NADP苹果酸酶
返回的是丙酮酸脱羧部位是线粒体
NAD苹果酸酶
返回的是丙氨酸脱羧部位是细胞质
PEP羧激酶
返回的是丙酮酸和丙氨酸㈢ CAM途径(景天科酸代谢途径)仙人掌,落地生根,菠萝,兰花等
气孔白天关闭,夜间气孔开放,固定二氧化碳。
夜间气孔开放 PEP+CO2 OAA+Pi
OAA 苹果酸 积累在液泡
白天气孔关闭 苹果酸 丙酮酸+CO2
PEP主要来自于呼吸作用的PPP途径和EMP途径。
这样,呼吸作用和光合作用相互联系起来。
调节
短期调节: 夜晚型和白天型
长期调节:兼性或诱导性CAM植物PEPC苹果酸脱氢酶苹果酸酶NADPNADPH发生在细胞质发生在叶绿体液泡叶绿体细胞质第三节 光呼吸 (Photorespiration)光呼吸
植物的绿色细胞在光下吸收O2,释放CO2的过程。
一、光呼吸的生化历程
光呼吸是一个氧化过程,被氧化的底物是乙醇酸。
㈠ 光呼吸底物乙醇酸的生物合成
RuBPC具有双重活性
RuBP + O2 2-磷酸乙醇酸 + 3-磷酸甘油酸
2-磷酸乙醇酸 乙醇酸 + 磷酸
以上是在叶绿体中进行的
㈡ 乙醇酸的氧化及CO2的产生
乙醇酸在叶绿体形成后,就转入另一个细胞器-过氧化物酶体。
C3植物叶肉细胞的过氧化物酶体较多, 而C4植物的过氧化物酶体较少,大多数在维管束鞘细胞的薄壁细胞内,一般位于叶绿体附近。 乙醇酸 + O2 乙醛酸 + H2O2
乙醛酸 + 谷氨酸 甘氨酸+α-酮戊二酸
甘氨酸转入线粒体,脱羧产生CO2
2甘氨酸 丝氨酸 + CO2 + NH3 + NADH2
丝氨酸回到过氧化物酶体,经转氨作用生成羟基丙酮酸
丝氨酸 + α-酮戊二酸 羟基丙酮酸 + 谷氨酸
羟基丙酮酸 + NADH2 甘油酸 + NAD+
甘油酸进入叶绿体,经磷酸化生成3-磷酸甘油酸,进入C3途
径,为进一步形成RuBP的光呼吸底物乙醇酸创造条件。
由于乙醇酸(底物)是C2化合物,氧化产物乙醛酸、甘氨酸
都是C2化合物,故也称光呼吸为二碳光呼吸氧化环或称C2环。 图3-26 光呼吸代谢途径线粒体 过氧化物酶体叶绿体磷酸乙醇酸NH3+CO2甘氨酸丝氨酸ATPADPNAD+NADH甘油酸二、光呼吸的特点⒈ 光呼吸随反应环境中氧浓度的增加而加速。
⒉ 光呼吸与光合作用关系密切。
⑴光呼吸底物乙醇酸来源于光合作用中的RuBP
⑵光合生成的有机物的1/3以上要消耗在光呼吸底物上
⑶光呼吸消耗能量
⒊光呼吸是在叶绿体、过氧化体、线粒体3 种细胞器联系作用下进行的。
⒋光呼吸的吸O2和排CO2不在同一个细胞器里,吸收O2是在叶绿体、过氧化体中,放出CO2在线粒体中。
三、光呼吸的生理意义
1. 防止强光对光合机构的破坏 光呼吸是一个保护性的生理过程,在高光强、低CO2 时会形成过量的同化力,由光激发的高能电子会传递给O2,形成超氧阴离子自由基会对光合膜、光合器官有伤害作用,而光呼吸可消耗同化力和高能电子,降低超氧阴离子自由基的形成,从而保护叶绿体,免除或减少强光对光合机构的破坏。
2. 维持C3循环的运转 在干旱和高辐射期间,气孔关闭,CO2不能进入,会导至光抑制,此时光呼吸释放CO2 ,能被C3途径再利用,避免产生光抑制。
3. 回收碳素 通过二碳氧化环可回收3/4的碳(2个乙醇酸转化1个PGA,释放1个CO2 ) 4. 对蛋白质合成提供了甘氨酸和丝氨酸。 5. 能消除乙醇酸的毒害作用。光呼吸可以转化乙醇酸。 6.可以为硝酸盐的还原提供还原型NAD四、比较C3植物和C4植物的光合特征 ㈠结构
㈡生理
⒈ 酶
⒉ CO2的补偿点
⒊ 光呼吸
⒋ 利用组织中CO2的情况
⒌ 热限
⒍ 所需能量
⒈酶对CO2的亲和力
PEPC 对CO2的Km = 7umol
RuBPC 对CO2的Km = 450umol
所以, PEPC对CO2的亲和力大。
⒉ CO2的补偿点
定义:指在照光情况下,植物光合作用所吸收的CO2量与呼吸作用所释放的CO2量达到动态平衡时外界环境中CO2浓度。
C3 植物CO2 补偿点高, C4 植物CO2 补偿点低。
⒊光呼吸
C3 植物是高光呼吸植物, C4 植物是低光呼吸植物
⑴ PEPC 对CO2亲和力要高于RuBPC
⑵ C4 植物RuBPC位于维管束鞘细胞,O2浓度较小;
⑶ 四碳二羧酸从叶肉细胞进入鞘细胞,起到“CO2泵”的作用,增加鞘细胞CO2浓度,增加RuBP 的羧化反应,减少了加氧反应,
⑷ C4植物PEP羧化酶位于叶肉细胞,对光呼吸释放的CO2 有捕捉作用
因此C4植物光呼吸小。
⒋利用组织中CO2的情况
C4植物①能利用细胞间隙中较低浓度的CO2
②光呼吸放出的CO2不易漏出。
⒌热限
C4植物热限比C3植物的热限高。
⒍所需能量
C4植物需额外的能量用于PEP的再生。
第四节 光合作用产物(photosynthetic product)主要是蔗糖和淀粉
一、淀粉的合成
(1)场所:叶绿体
(2)原料:来自于C3途径的磷酸丙糖
DHAP+PGAld FBP F6P G6P G1P
G1P+ATP ADPG
( G)n
二、蔗糖的合成
(1)场所:细胞质
(2)原料:来自于C3途径的磷酸丙糖
(3)磷酸运送器(Phosphate translocator)
磷酸运送器(Phosphate translocator)
是在叶绿体和胞质溶胶之间转运磷酸的专一载体。每从叶绿体运出一个磷酸酯就有一个正磷酸离子从胞质溶胶运送回来。
磷酸丙糖 + Pi 磷酸丙糖 + Pi
(叶绿体) (细胞质) (细胞质) (叶绿体)
DHAP+PGAld FBP F6P G6P G1P
G1P+UTP UDPG
F6P
三、蔗糖和淀粉合成的调节
由磷酸和磷酸丙糖的相对浓度控制。
Pi, F6P促进蔗糖合成。磷酸运送器蔗糖-6-磷酸蔗糖合酶蔗糖第五节 影响光合作用的因素一、光合作用的指标
1、光合强度(Intensity of photosynthesis)
也称光合速率 (Photosynthetic rate)
单位时间、单位叶面积吸收CO2的量或放出O2的量。
2、净光合强度(表观光合强度)
实际所测得的光合速率
半叶法、红外CO2分析仪法、氧电极法
3、总光合速率(Gross Photosynthetic rate )
也称真光合速率(Ture Photosynthetic rate )
表观光合速率加上光、暗呼吸速率
二、影响光合作用的因素1、光照 : 光强、光质
2、CO2浓度
3、矿质营养
4、水分
5、温度:三基点温度
6、光合作用的日变化1、光照 : 光强、光质
⑴光补偿点(Light compensation point)
同一叶子在同一时间内,光合过程中吸收的CO2和呼吸过程中放出的CO2等量时的光照强度。
⑵光饱和现象(Light saturation) /光饱和点(Light saturation point)
当达到某一光强时,光合速率就不再随光强的增强而增加,这种现象称为光饱和现象。
刚刚出现光饱和现象时的光照强度称为光饱和点。
⑶光抑制(Photoinhibition)
光能超过光合系统所能利用的时,光合功能下降的现象叫~。
⑷ 光质:红光最有效2、CO2浓度
⑴ CO2饱和点(CO2 Saturation point)
在一定范围内,植物净光合速率随CO2浓度升高而增加,但到达一定程度时,再增加CO2浓度,净光合速率不再增加,这时CO2浓度叫CO2饱和点。
⑵ CO2补偿点(CO2 Compensation point)
降低环境温度可以降低光补偿点。(??? ) CO2的补偿点随光照强度的变化而变化。(??? )CO2饱和度随光照强度的增加而升高.(??? ) 3、矿质营养
直接、间接影响光合作用
调节气孔开度,影响碳水化合物的转换及运输,光合链成分,叶绿素组分,酶的活化剂,加大光合面积等。4、水分
⑴ 缺水引起气孔开度小或关闭,限制了CO2的吸收----气孔阻力
⑵ 缺水时,淀粉水解加强,糖类堆积,光合产物输出缓慢,抑制光合作用的进行,而促进了呼吸作用。
(3)缺水减少光合面积。
(4)缺水会降低CO2在细胞间隙内的传递速度---叶肉阻力。
5、温度:三基点温度
C4植物的最适温度高于C3植物
C4植物的热限高于C3植物
6、光合作用的日变化
⑴ 温暖的日子,水分充足,太阳光是主要矛盾,光合速率与太阳辐射进程相符合。
⑵ 如白天云量变化不定,光合速率呈不规则曲线。
⑶晴天无云,但光强强烈时,“午休”现象。
光合“午休”现象(midday depression)
指植物的光合速率在中午前后下降的现象。引起“午休”的主要原因是大气干旱和土壤干旱及强光、高温、低CO2浓度等。
第六节 植物对光能的利用一、光能利用率
指植物光合作用所积累的有机物所含的能量,占照射在单位地面上的日光能的比率。一般为5%。
1、量子需要量:每同化1分子CO2或释放 1分子O2所需的光量子数。
2、量子产额或量子效率:每吸收1个光量子所能同化的CO2或释放 的O2的分子数。
3、生物学产量:光合产量-光合产物消耗。
4、光合产量:光合面积×光合强度×光合时间
5、经济产量:生物学产量×经济系数
6、经济系数:指收获物中经济价值较高的部位的产量占作物总
量的比值。
光能利用率低的原因: 不能吸收的波长 60%,
反射和透光 8%
热能散失 8%
代谢 19%
转化 5%二、光合产物的分配规律
由“代谢源”到“代谢库”
代谢源:指制造和输出同化物的部位,如成熟的叶片。
代谢库:指输出和利用同化物的部位,如果实、种子。
⒈就近供应
⒉优先供应生长中心
⒊同侧供应
提高植物光能利用率的途径
经济产量=(光合面积×光合速率×光合时间-消耗)×经济系数
延长光照时间
提高复种指数(全年内作物收获面积与耕地面积之比):套种或间作。
延长生长期:如育苗移栽、套种、适时早播、防止早衰。
补充人工光照
增加光合面积
合理密植
改善株型:杆矮、叶直而小、叶片厚、分蘖密集。
加强光合效率
提高CO2浓度:通风、施CO2(干冰)、施有机肥、碳铵
抑制光呼吸:光呼吸抑制剂,如α-羟基磺酸可抑制乙醇酸氧化酶活性。
提高经济系数(经济产量与生物产量之比)测试1、在光照强,温度高,空气湿度低的情况下, C3 和C4植物哪一个光合效率高?为什么?(光饱和点,热限,CO2泵, CO2补偿点, CO2亲和力,光呼吸,利用组织CO2情况)
2、在光照弱,温度低,空气湿度高的情况下, C3 和C4植物哪一个光合效率高?为什么?
3、在碳同化中受光调节的酶有哪几种?
4、如何证明光合作用中释放的氧气来自水。
5、如何证明光合电子传递有两个光系统参与?
6、在缺乏CO2的情况下,对绿色叶片照光能观察到荧光,然后在供给CO2的情况下,荧光立即被淬灭,试解释其原因。
补充:叶绿素的理化性质⑴ 荧光现象和磷光现象
⑵ 皂化现象
⑶H,Cu代镁现象
+2KOH+ CH3OH+C20H39OH补充:叶绿素的层析
胡萝卜素叶黄素叶绿素a叶绿素b