第四章 种群
图片预览











文档简介
课件175张PPT。种群生态学海葵 种群术语起源于人口统计学。其英文“population”自拉丁语词根“populus”,原词义是“人”或“人民” (people),因此被译为“人口”。早期的生态学者在研究其他生物时,如昆虫、鱼类、鸟类等,曾经将它们相应地翻译为“虫口”、“鱼口”、“鸟口”等。台湾、香港或海外的华人也有的将“population”翻译为“族群”,或“繁群”。目前,我国的生物学者已一致将“population”翻译为“种群”。种群可以再分为繁殖群(deme)或地方种群(local population),即一群能够相互交配繁殖的同种个体。 种群生态学种群是一定区域内同种生物个体的集合。种群间的边界可以是任意的。种群可以根据组成种群的生物是单体生物还是构件生物进行分类。在单体生物种群中,每一受精卵发育成一单个个体。在构件生物种群中,受精卵发育成一个结构单位,这一结构单位再形成更多的构件和分支结构。然后这些结构可能分裂,形成许多无性系分株。种 群种群生态学种群生态学海葵种群茅莓种群种群生态学肯尼亚的斑马种群
种群生态学是研究种群生物系统的规律的科学,具体地说,就是研究种群内部各成员之间,种群(或其成员)与其他生物种群之间,以及种群与周围环境非生物因素的相互作用规律。
种群生态学的重要内容之一是种群动态研究。种群动态研究也就是研究种群数量在时间上和空间上的变动规律及其变动原因。种群生态学 种群数量决定着该种群对生态系统作用的大小,对于有害动物或杂草、经济动物或植物、或是濒危物种,则相应反映着它们的危害程度、可利用程度以及濒危程度。
种群并不是简单的相加,种群作为一种新的生物系统应该有新质产生。种群生态学银杏 第一节 种群密度和分布
不同生物学分支学科对种群这一概念的理解存在着差异。
生态学上的种群概念强调了种群的分布和数量,并且强调了种群是一种生物系统,即种群不仅与环境相互关联,而且种群内部个体之间存在着遗传信息交换等种内关系。
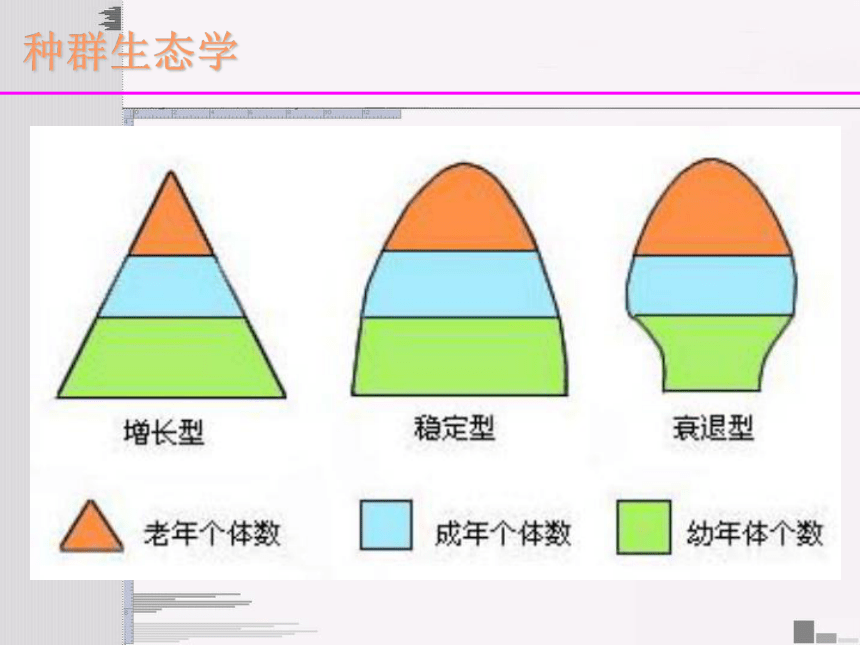
种群生态学种群生态学种群密度很高种群密度一般种群生态学对于单体生物和种群如哺乳类,其种群大小就是一定区域内个体的数量,非常简单。对于构件生物,如植物和珊瑚,情况就较复杂。对于这些种群,“断片”(无性系分株)或枝条(构件)的数目比不同个体的数量更有意义,代表多度。种群大小种群生态学种群生态学种群的年龄结构是每一年龄阶段个体数目的比率,通常以年龄金字塔图来表示。既不增长也不下降的种群有稳定的年龄分布。增长型的种群有更多的年轻个体,而在下降型种群中年老的个体占优势。当种群经历离散的发育时期(如昆虫幼体的龄期)时,每一时期个体的数目(“时期结构”)可以对种群进行有效的描述。对于生长率无法预测的物种(如植物),根据大小分类可能更有意义。年龄和时期
结构 年龄锥体的三种基本类型
A增长型种群: 幼年组个体数多,老年组个体数少,种群的死亡率小于出生率,种群迅速增长。
B稳定型种群: 种群出生率大约与死亡率相当,种群稳。
C下降型种群: 幼年组个体数少,老年组个体数多,种群的死亡率大于出生率,种群种群数量趋向减少。繁殖后期繁殖期繁殖前期ABC种群生态学稳定年龄分布(stable age distribution)
年龄金字塔(age pyramid)
固定年龄分布(stationary age distribution)
植物体作为构件生物,有年龄结构,由年轻的、发育中的、正在起作用的和衰老的部分组成。企鹅种群生态学不同构件的植物种群生态学出生率就是新个体的产生,实际出生率就是一段时间内每个雌体实际的成功繁殖量。特定年龄出生率就是特定年龄组内雌体在单位时间内产生的后代数量。出生率死亡率是在一定时间段内死亡个体的数量除以该时间段内种群的平均大小。这是一个瞬时率,可用来估价整个种群的死亡率或特定年龄群的特定年龄死亡率,死亡的概率是死亡个体数除以在每一时间段开始时的个体数。死亡率种群生态学迁入率(immigration):进入当前种群领地的同种个体占总个体数的比率;
迁出率(emigration):离开当前种群领地的同种个体占总个体数的比率。
A型——凸型(convex curve)的存活曲线。 种群生态学B型——对角线型(diagonal straight 1ine)的存活曲线。 种群生态学C型——凹型(concave curve)的存活曲线。 种群生态学种群生态学存活率是死亡率的倒数。对于一个特定种群,存活率的数据通常以存活曲线的形式来表示;存活曲线表示的是在每一个生活期存活个体所占的比率的对数值。根据各种生活期死亡率的高低,特定年龄存活曲线一般有三种模式:后期死亡率最高(类型I),各期死亡率相等(类型II),早期死亡率最高(类型III)。存活率种群生态学树袋熊考拉种群大小随时间的变化可以按如下方法计算:t时间种群原来数量(Nt),加上新出生的个体数(B)和迁入个体数(I),减去死亡个体数(D)和迁出的个体数(E),就可得到t+1时间种群的数量(N t+1),这可用以下方程表示。
N t+1= Nt +B + I – D – E
在一组特定条件下,一个体具有最大的生殖潜力,称为内禀自然增长率r。这是种群在不受资源限制的情况下,于一定环境中可达到的理论最大值。种群生态学种群增长种群生态学种群生态学逻辑斯谛方程描述的是一个在有限资源空间中的简单种群的增长。在早期,资源丰富,死亡率最小,繁殖尽可能的快,种群内个体可达到内禀增长率。种群呈几何式增长,直到种群数量达到环境可持续支持的最大程度,这个最大数量称为环境容纳量(K)。当种群更加拥挤时,种群增长率减少到零,种群大小处于稳定状态。这可用逻辑斯谛方程来表示:
t时间种群大小变化率=内禀增长率×种群大小×密度制约因子
dN/dt = rN(1 - (N/K))
当种群达到环境容纳量,种内竞争变得更激烈时,密度制约因子(1 - (N/K))会接近零。该方程预测种群的增长随时间变化呈现出“S”形,如在真实种群中通常所观察的那样。密度制约性
种群增长:
逻辑斯谛方程种群的内禀增长率(intrinsic rate of increase):
自然增长率(rate of natural increase)是种群的实际增长率,用r来表示。它是指在单位时间内某一种群的增长百分比。
增长率=自然增长率+净迁移率=出生率-死亡率+迁入率-迁出率
内禀增长率或称为内在增长率(rm),即在无限制的环境条件下,种群的最大增长率。内禀增长率也称为瞬时增长率(instantaneous rate),或称为生物潜能(biotic potential)或生殖潜能(reproductive potential)。种群生态学苏格兰松和领鸽种群的指数增长曲线种群生态学 逻辑斯谛曲线可分为5个部分,即种群增长的5个时期:?
(1)开始期:潜伏期或缓慢期,初始种群刚刚适应环境。
(2)加速期:当初始种群适应环境后,个体加速繁殖,种群迅速增长,增长速度取决于初始种群的数量(N0)和瞬时增长率(r)。 ((3)转折期:种群增长达到最大后开始逐渐地减少。
(4)减速期:种群在这一时期的瞬时增长率迅速地下降。
((5)饱和期:种群密度接近或达到k值,不再继续增长。种群生态学 逻辑斯谛增长模型的意义:
(1)是许多两个相互作用种群增长模型的基础;
(2)是农、林、渔业等实践领域中,确定最大持续产量(maximum sustainable yield)的主要模型;
(3)模型中两个参数r和K已成为生物进化对策理论中的重要概念。生态密度(ecological density)单位面积(或体积)特定生态环境中的生物个体数。
根据密度调查方法的不同,密度可以分为绝对密度(absolute density)和相对密度(relative density)。种群生态学(一)绝对密度的常用调查方法
1.总数量调查法(total count)。又称直接统计法,是直接计数一定地方的所有个体,由此得到种群密度。E.g.:人口调查法(census technique)
2.取样调查法(sampling methods)。大多数绝对密度调查只是在几个地方计数种群的一小部分,由此估计种群整体的密度。
常用的取样调查法有三种:
①样方法(use of quadrate)
②标志重捕法(mark recapture methods):N=(M×n)/m
M是标志数,n是再捕个体数,m再捕中标记数
③去除取样法(removal sampling) :对于一个封闭的种群,对逐次捕获数/单位努力(作为y轴),逐次捕捉的累计数(作为x轴)作图,就可以得到一个回归线,到达 x轴的截距所表示的数量就是种群数量N。种群生态学(二)相对密度的调查方法
1.动物计数(counts of animals)
2.动物痕迹的计数(counts of animal signs)
3.单位努力捕获量(catch per unit effort)。
每一种生物的种群密度,都有一定的变化限度。
种群生态学白鹭计数时以其痕迹为依据 最大密度(maximum density)或称饱和密度(saturation density或upper asymptote level)是指特定环境所能容纳某种生物的最大个体数。当超过这一密度,种群数量将不再增长。
最小密度(minimum density或lower asymptote level)是指种群维持正常繁殖、弥补死亡个体所需要的最小个体数。如果低于最小密度,由于难以寻找配偶。繁殖意愿丧失、繁殖力下降,结果,种群就难以生存。
在最大密度和最小密度之间存在着一个最适密度(optimum density),当处于最适密度状态时,种群的增长最快。种群生态学处于饱和密度的森林种群生态学川金丝猴雪豹穿山甲犀鸟濒危动物性比率(sex ratio)是指种群中雄性与雌性个体数的比例。
大多数动物种群的性比接近1:1;
营社会生活的昆虫雄性多于雌性;
孤雌生殖的轮虫和枝角类等雌性多;
还有随环境条件的变化而性别改变的,水生动物如虾、黄鳝等。 种群生态学种群生态学轮虫河虾沼虾黄鳝种群生态学如果种群参数如出生率和死亡率随着种群密度的变化而变化,就可以说是密度制约的。如果出生率和死亡率不随着密度的变化而变化,则是非密度制约性的。如果没有迁移,除非出生率和死亡率是密度制约性的,否则种群会持续增长。
密度制约种群生态学种群生态学密度制约出生率和死亡率的平衡给出平衡密度,或环境容纳量 K。 (a)出生率和死亡率都是密度制约的;(b)非密度制约出生率;
(c)非密度制约死亡率;(d)同(a),但出生率和死亡率结合环境变化。种群生态学平衡种群密度当单位个体出生率正好平衡单位个体死亡率,种群密度既不增加也不减少时,此时的种群密度为平衡种群密度。平衡种群密度与环境容纳量K值相等。目前趋于平衡的斑马种群生态学密度制约有三种形式:过度补偿,补偿不足和准确补偿。如果是过度补偿,当初始密度上升时,存活个体数目将下降。如果密度制约是补偿不足,当初始密度上升时,存活个体数量上升。如果下降个体数与开始增加的个体数相等,则称密度制约为准确补偿。补偿和密度
制约表明的是密度制约性过程与种群对可利用资源的利用的对抗关系种群生态学大多数实际种群并不是在平衡密度停留很长时间,而是动态的不断的发生变化。由于周围环境的变化或生物因子的影响,种群可能会增长或收缩。增长种群和
收缩种群迁徙耕作种群生态学种群生态学种群生态学种群生态学混 沌数学上有关混沌的定义与俗语中的用法是截然不同的。一个混沌系统是受确定性作用控制的,而不是随机的。混沌的结果依赖于初始状态的精确值。由于完全精确是不可能达到的,因此对混沌系统不能进行有效的预测。一些分析认为,麻疹的爆发和一些昆虫种群可能表现出混沌的动态变化。种群生态学蟋蟀螳螂黑甲虫卵麻疹易于混沌的昆虫种群生态学季节消长:季节变动引起的自然种群数量变动,如北点地梅
种群爆发:具有不规则或周期性波动的生物都可能出现种群爆发,如害虫、赤潮等。
生态入侵(ecological invasion):由于人类有意识或无意识地把某种生物带入其适宜其栖息和繁衍的地区,该生物种群不断扩大,分布区逐步稳定地扩展,这种过程为生态入侵。水葫芦种群生态学蝗虫种群爆发种群生态学澳大利亚的侵入种群种群生态学种群的分布格局
组成种群的多个个体在其生活空间中的位置或布局,称为种群的内分布型(internal distribution pattern) :
(1)集群型(clumped)
(2)随机型(random)
(3)均匀型(uniform)树蕨,一种只分布于四川的植物集群型随机型均匀型种群生态学通常用泊松分布、负二项分布、奈曼分布等模型计算 第二节 种群的相互作用关系
一、集群
集群(aggregation或society, colony) 现象普遍存在于自然种群中。同一种生物的不同个体,或多或少都会在一定的时期内生活在一起,从而保证种群的生存和正常繁殖,因此集群是一种重要的适应性特征。
根据集群后群体持续时间的长短,可以把集群分为临时性(temporary)和永久性(permanent)两种类型。 种群生态学 生物产生集群的原因是复杂多样的,包括:
⑴对栖息地的食物、光照、温度、水等生态因子的共同需求;
⑵对昼夜天气或季节气候的共同反应。如过夜、迁徒、冬眠等群体;
⑶繁殖的结果;
⑷被动运送的结果;
⑸由于个体之间社会吸引力(social attraction)相互吸引的结果。
动物群体的形成可能是完全由环境因素所决定的,也可能是由社会吸引力(social attraction)所引起。种群生态学 集群的生态学意义:
(1)集群有利于提高捕食效率;
(2)集群可以共同防御敌害;
(3)集群有利于改变小生境;
(4)集群有利某些动物种类提高学习效率;
(5)集群能够促进繁殖。
拥挤效应(overcrowding effect)种群生态学种群生态学相互作用的
分类个体或物种间的相互作用可以相互作用的机制和影响为基础来分类。关键的种间相互作用是竞争、捕食、寄生和互利共生,而主要的种内相互作用是竞争、自相残杀和利他主义。偏利共生(commensualism):如寄居蟹与水螅虫,对后者有利,对前者无害;
偏害共生(amensualism):有毒物种和土壤真菌之间;
拟寄生(parasitoidism):通常指昆虫。种群生态学竞 争竞争是共同利用有限资源的个体间的相互作用,会降低竞争个体间的适合度。竞争即可在利用共同资源的物种间发生(种间竞争),也可在同种个体间发生(种内竞争)。个体或物种的生态位(它所处的环境,利用的资源和它发生的时间)是决定该个体或物种与其他个体或物种竞争程度的关键。大范围的生态位重叠一般导致激烈竞争。种群生态学当个体对资源的需要非常相似时,竞争会特别强烈。种内竞争是生态学的一种主要影响力,是扩散和领域现象的原因,并且是种群通过密度制约过程进行调节的主要原因。
如欧鳊,一种淡水鱼,幼鱼摄食小的浮游动物,成鱼以大型底栖无脊椎动物为食。种内竞争成鱼种群生态学种间竞争种间竞争发生在利用同样有限资源的两物种之间。极少种能够逃脱其他种与之竞争共同资源的影响。
如双核小草履虫和大草履虫在酵母介质中培养,结果前者占优胜,后者死亡等。种群生态学竞争有两种作用方式。在资源利用性竞争方式下,个体不直接相互作用,而是耗尽资源使供应不足。由于可利用资源不足而造成适合度下降。利用性竞争种群生态学在干扰性竞争方式下,个体直接相互作用,在一些动物种类中,最明显的通过打斗,也通过产生毒物(如植物异株克生)进行竞争。在这种相互作用中“败者”适合度下降,可能由于干扰(如受伤或死亡),也或许由于缺乏可用资源。干扰性竞争种群生态学他感作用:通常指一种植物通过向体外分泌代谢过程中的化学物质,对其他植物产生直接或间接的影响。许多高等植物能产生具有自毒(autotoxic)和抗生性质的有机物质,如碳水化合物中的萜烯、酒精、有机酸、醚、醛、酮等类物质。这些有机物质从各种途径释放出来,如根分泌直接进入土壤、从活或死组织淋洗出来、或挥发到空气中散布开来。这些物质能够抑制或促进其他物质的生长。
如:Encelia farinosa 一种菊科植物,叶子分泌的一种苯甲醛物质对相邻的番茄、玉米等物质有强烈的抑制作用;风信子、稠李、洋槐、丁香等的花,熏衣草、薄荷、月桂等的叶子的芳香物质对相邻植物伸长生长起抑制作用;榆属与栎树、白蜡树与栎树、白桦与松树、松树与云杉不能混种在一起;胡桃分泌物毒害苹果,对番茄和马铃薯也有影响;玉米对苹果有抑制作用;葱和菜豆、芜菁和番茄、番茄和黄瓜、冬黑麦和小麦之间都有抑制作用。
另一些植物,如洋葱和食用甜菜、马铃薯和菜豆、小麦和豌豆生长在一起却能彼此促进。正在猎食中的狮子领鸽(Streptopelia decaocto)的种群扩散过程自M.C.Molles,Jr,1999 非洲蜂(Apis melifera)的种群扩散自M.C.Molles,Jr,1999北美的枫树(Acer spp.)(左)和芹叶钩吻(Tsuga canadensis)(右)的种群扩散自M.C.Molles,Jr,1999种群生态学许多动物种(包括昆虫、鸟类和哺乳动物)的个体或群体为争夺空间而竞争。保持领域边界的个体间有积极的相互干扰行为。领域使其所有者受益,为对抗入侵者,保护领域所付出的代价,被增加的食物供应、提高的交配成功率和降低的被捕食的危险所带来的价值超过。领 域 性社会等级(social hierarchy):动物种群中的支配和从属关系。保卫“我”的领地种群生态学固着生长的生物,包括植物,不能通过运动逃避竞争,因此竞争中的失败者死去在同样年龄大小的植物群中,这种竞争结果使较少量的较大个体存活下来。这一过程叫做“自疏”。自疏导致密度与植物个体大小之间的关系,这在双对数作图时呈现典型的-3/2斜率。这种关系叫做Yoda-3/2自疏法则。自 疏种群生态学菊花的自疏种群生态学如果两个物种在稳定环境中竞争,则有两种可能的结果:(i)一种被排除,或(ii)两种共存。竞争排斥原理陈述,共存只能发生在两物种生态位分化的稳定环境中。竞争排斥种群生态学In the absence of a competitor, a species may expand its niche. Examples of such competitive release include ground doves in New Guinea and gerbils in Israel.Competitive release种群生态学在缺乏竞争者时,物种会扩张其实际生态位。这种竞争释放的例子包括新几内亚岛上的地鸽子和以色列的沙鼠。竞争释放种群生态学当明确的生态位契约受到竞争影响时,为了适应新的资源谱,个体形态会相应变化。这种性状替换发现在收获蚁Veromessor Pergandei 和达尔文雀Geospiza fortis和G. fuliginosa。性状替换种群生态学环境是许多不同质量、不同资源水平的生境的拼缀。一些斑块的发生是暂时的和不可预测的。在时间和空间上,环境是异质性的。这意味着竞争“战斗”在环境变化和竞争平衡变化之前不会完成。因此,在现实世界中共存的种类,其共存可能是因为环境的异质性,而不是因为生态位分化。空间和时间
异质性种群生态学如果捕食者进攻两种猎物,那么每一种猎物,会通过增加当地捕食者种群的数量,相反地影响另一种。因此,两个猎物种群之间的相互作用看上去似乎它们确实是在竞争,然而它们或许利用完全不同的资源。这一现象叫做表观竞争。表观竞争种群生态学五、捕食种群生态学捕食可定义为摄取其他个体(猎物)的全部或部分为食。这一广泛的定义包括
(i)典型的捕食者,在袭击猎物后迅速杀死而食之;
(ii)草食者,只消费对象个体的一部分;(iii)寄生者,与单一对象个体(寄生)有密切关系,通常生活在寄主的组织中。捕食的定义等待猎物的老虎种群生态学捕食者可以划分为(i)消费植物组织的草食者,(ii)摄食动物组织的肉食者和(iii)既摄食植物组织也摄食动物组织的杂食者。动物、植物食性的差异需要不同的生理和行为适应,从而导致肉食者与草食者世系之间反复的进行分歧。肉食者和
草食者种群生态学捕食者随其摄取猎物的数量多少而变化,某些捕食者是特化种,而另一些是泛化种。一般来说,寄生者比典型捕食者更为特化、而草食者比肉食者更为特化。泛化种和
特化种种群生态学种群生态学捕食者和寄生者是否能够调节其猎物种群的大小呢?这一问题并不像看上去的那么简单。有两种主要观点:(i)任一捕食者的作用,只占猎物种总死亡率的很小一部分,因此去除捕食者对猎物种仅有微弱效果;(ii)捕食者只是杀死了对象种中即将死亡的个体,所以最终对猎物种群大小没有影响。然而,在一些例子中确有明显证据表明捕食者对猎物数量有重要影响。捕食者对猎物
种群大小的
影响种群生态学种群生态学捕食者-猎物模型行为多度捕食者猎物时间N种群生态学捕食者和猎物的种群动态分为4个时期:a猎物增加,捕食者减少;b猎物和捕食者都增加;c猎物减少,捕食者继续增加;d猎物和捕食者都减少。种群生态学猎物收益率种群生态学猎物转换依据猎物种数量的多少,捕食者有时会转换其选择而捕食某一特定的猎物种。这时捕食者主要捕食优势种猎物而大大忽略其他猎物。猎物密度影响
——功能反应一般认为在高猎物密度下,捕食者的摄食率会增加,然后随猎物饱和达到最大速度。这种关系称为功能反应,可能采用多种模式,传统上分为三种类型:功能反应I、II和III。种群生态学Ⅰ随猎物密度的增加,捕食者的摄食以固定比例增加;Ⅱ捕食者在摄食猎物时可能达到饱足,摄食率降低,直到完全饱足,摄食率不再随猎物密度的变化而变化;Ⅲ捕食者在猎物密度低时由于低狩猎效率而反应低,以后摄食率又因饱足水平而减少。种群生态学搜寻和处理为得到食物,捕食者必须首先搜寻猎物,然后处理(抓住、加工和吃掉猎物)。可以认为捕食者食谱的宽度是由泛化种对策与特化种对策之间的权衡决定的。泛化种对策捕食者寻找多种猎物(相对容易),特化种对策捕食者寻找一类猎物,非常有效的处理它。最佳觅食理论假定进化会最优化动物行为以使其获得的能量效率最大,从而做出捕食者如何权衡搜寻与处理的预测。种群生态学狩猎者 种群生态学种群生态学种群生态学疾藜草─是兇惡植
物,身上插滿一根根
堅硬又尖銳的針刺。
任何動物只要一不小
心惹上它們就會疼痛
不已。种群生态学寄生物的
多样性寄生物是摄取其他生物的组织、与其寄主紧密相联而生的捕食者的子群。寄生物可以分为两大类(i)微寄生物,在寄主体内或表面繁殖,(ii)大寄生物,在寄主体内或表面生长,但不繁殖。主要的微寄生物有病毒、细菌、真菌和原生动物。寄生蠕虫和昆虫是重要的大寄生物。一大类昆虫大寄生物是拟寄生生物,它们在昆虫寄主身上或体内产卵,通常导致寄主死亡。鸡肠中的蛔虫种群生态学种群生态学种群生态学SARS病毒种群生态学传播方式寄生物可以横向传播(在种群个体之间),或在少数情况下,纵向传播(从母体到后代)。横向传播或直接或间接,由传病媒介(如蚊子)或中间寄主做中介。种群生态学寄主对疾病的反应脊椎动物被微寄生物感染后产生强烈的免疫反应。这种反应主要有两种成分:(i)细胞免疫反应,特定细胞直接攻击病原体细胞,和(ii)B-细胞免疫反应,使抗体量上升。初次被病原体感染后,免疫记忆对未来同样的侵袭产生快速反应,导致免疫。无脊椎动物和植物也可以抑制感染,但方式较简单,通常较少特化机制。免疫造成的潜在寄主减少,使微寄生物的繁荣-和-破产对策加强.种群生态学疾病的传播途径种群生态学种群生态学社会性寄生物:“社会性寄生物”与其寄主之间有一种完全不同的关系。社会性寄生物不通过摄食寄主组织获益,而是通过强迫寄主提供食物或其他利益而获利。这种关系发现在杜鹃,它们将蛋下在别种鸟的巢中,让这种鸟饲育其幼鸟。这种关系还发现在一些蚂蚁种类,它们迫使其他种工蚁给它们的幼体食品或其他利益。
种群生态学工蚁种群生态学种群生态学种群生态学蜜蜂与花的协同进化种群生态学寄主——
寄生物进化寄生物与其寄主间紧密的关联经常导致二者在进化上的相互作用,或协同进化。协同进化使寄主的防御机制提高,而给寄生物克服这些防御的通路——这叫做“军备竞赛”。但是,寄主与寄生物之间的协同进化并不都是提高性的。例如,协同进化会降低寄生物的毒性。 大熊猫体内存有纤毛虫种群生态学寄生物种群的异质性一些疾病是稳定的,而另一些进化了通过变异“逃离”寄主防御的手段。人类“孩童时期”的疾病——麻疹、腮腺炎、水痘等大部分局限于孩子,因为它们的抗原稳定。与此相对,寄生物如流感病毒、鼻病毒(普通感冒原)和沙门氏菌(Salmonella)具有可经常加入的很多变异型。寄主因此要经常与变化的寄生物军队的进攻而进行战斗。种群生态学种群生态学七、互利共生( Mutualism)互利共生和
共生现象互利共生是不同种两个体间一种正的互惠关系,可增加双方的适合度。互利共生可以是共生性的,生物体以一种紧密的物理关系生活在一起。种群生态学地 衣种群生态学蝴蝶传粉种群生态学种群生态学传 粉异型杂交植物需要将其花粉转移到同种植物的柱头上,并接受同种植物的花粉。一些植物种类靠风来传播花粉,如果植物生长在广大的植物种类单一的地方如草地和松林,这种传播方式会很有效。但是大多数开花的双子叶植物,依靠昆虫、鸟、蝙蝠或小型哺乳动物来传播花粉,作为交换,这些动物或以花蜜或以花粉本身为食。种群生态学传 粉种群生态学两种蜂鸟及其特化对应的花的形态种群生态学种子扩散大型种子无法有效的被风传布,除非靠水传布(如椰树的种子),或依靠动物来散布。啮齿动物、蝙蝠、鸟和蚂蚁都是重要的种子扩散者。
食水果者(frugivore)来自澳洲的老鼠种群生态学浦公英种群生态学互利共生和
进化互利共生可能起源于寄生物-寄主和捕食者-猎物之间的关系,或起源于没有协作或相互利益的紧密共栖物种之间。然后双方的进化变化(协同进化)导致双方从这种关系中获益,虽然也可能共生“恶化”,成为使一方受益、另一方受非平衡的剥削的寄生关系。互利共生的相互作用在多细胞动物进化过程的一些关键阶段中起着中心作用。高等生物的许多细胞器官,包括线粒体和叶绿体(都来自自由生活的原核生物,含有环状DNA基因组和其他细胞特征),被认为来自共生性细胞。互利共生的例子:人与家畜(不能独立),丝暇虎鱼利用鼓虾的巢穴作隐蔽场所,鼓虾(不能独立)是盲的,离开洞穴时,以细长的触角保持与鱼的接触,代替眼睛起警报作用。后者如花和蜜蜂,反刍动物与瘤胃内的细菌和原生动物,高等植物与真菌(菌根),白蚁和肠道内的鞭毛虫等。
红豆杉大量繁殖的竹子种群生态学 迁移:生物也可以通过移动到别处来躲避当地恶劣的环境。迁徙是方向性运动,如燕子从欧洲到非洲的秋季飞行。相反,扩散是离开出生或繁殖地的非方向性运动。迁移有三种,依据生物个体做的是(i)反复的往返旅行,(ii)单次往返旅行(死之前回道出生地进行繁殖),或(iii)单程旅行(一个世代向一个方向运动,下一个世代返回来)。
迁徙中的鸟种群生态学种群生态学种群生态学复杂的生活
周期许多植物、真菌和动物具有复杂的生活周期,在生活周期中,或者个体采用根本不同的形态学性状(如毛虫/蝴蝶,蝌蚪/青蛙)或者以一种可预测的方式采取彼此根本不同的世代(如植物的单倍体/双倍体世代交替,锈菌和蚜虫的有性/无性世代交替)。为什么要采用这些复杂的生活史对策呢?在一些种类、不同的时期,或者主要进行生长,或者主要进行扩散,而在其他种类,不同的时期是为了最有效地利用不同的生境。种群生态学衰 老为什么生物个体变老后身体恶化,繁殖力和精力降低?对这一问题有两个水平的答案。在机制水平上,由于毒物和自然辐射的影响,使细胞机器崩溃,从而引起衰老。但是,这不能是全部的原因——衰老的发生随种类不同变化很大。这说明进化影响或决定着衰老。衰老的进化模型认为,或者(i)一个影响较老个体的有害突变,比影响年轻个体的有害突变,受到更弱的选择对抗,或者(ii)对早期繁殖有益的基因在生命晚期有恶性影响。
又称为突变积累(mutation-accumulation),早期表达的基因会显著降低个体的繁殖输出;拮抗性多效(antagnistic pleiotropy),如马鹿的一个与提早繁殖和高繁殖力相关的基因也与低存活力有关,而黑尾果蝇对提高的存活力的选择使早期繁殖力降低。默契的天鹅种群生态学合作的蚂蚁种群生态学加里福尼亚鸟群蜂王种群生态学生态学中的性性性是植物和动物中广泛分布的现象,性有一系列生态后果。性在进化上的重要性是由于它产生遗传变异,它在生态学上的重要性则是因为不同性别的生物其行为不同。两性之间的区别是由于它们对后代发育的能量投资不对称性而产生的。(很多雄性仅提供精子或花粉,雌性却要提供大的卵,子宫胚珠和乳汁等)种群生态学近交的代价当亲缘相近的个体进行近亲繁殖时,“近交”就出现了。近交引起(i)近交衰退(它使后代的适合度降低)和(ii)增加遗传纯合性(它降低了对于环境变化产生进化适应的可能性,而增加了灭绝的可能性)。(有害隐性等位基因的纯合频率的增加在亲缘关系近的种群中更容易出现,是近交衰退的原因。)许多物种具有防止或减少近交的机制。笼养动物和野生动物种群生态学这是全球发现的第一只饲养中雌雄同体的蝴蝶。这只大凤蝶最近从新加坡圣陶沙的蝴蝶园内破蛹而出。种群生态学交配体制动物,特别是脊椎动物,其交配的社会结构是重要的生态特征。一个雄体和一个雌体形成配偶,或者仅在繁殖季节,或者直到其中有一个死亡,这是单配制(monogyny)(鸟类)。当一个雄体与多个雌体交配,就出现了一雄多雌制(polygyny)(哺乳类);当出现相反情况(很少见),即一个雌体具有多个雄体的交配群,就叫做一雌多雄制(polyandry)(北极冻原的鸻科鸟和最严酷的喜马拉雅地区的人类社会)。种群生态学种群生态学性 比性比是种群中雄体对雌体的比率,通常接近于1雄:1雌。但是,不等的消耗(条蜂,雌性比雄性重58%,理论值性比为1.58:1,实测值为1.63:1)、拥挤的配偶竞争(雌体间的局域资源竞争紧张,产出的雄体离家出走)也可能使性比偏离。种群生态学在孔雀的种群中,雄性个体比雌性多得多种群生态学性 选 择性选择导致形成像孔雀之尾、雄苇莺动听的歌唱、雄鹿的叉角等特征。交配竞争中生殖成效的不同导致性选择。
有性内选择(打斗)和性间选择(偏爱异性的某个独特特征。让步赛理论(handicap theory)即好性状有好基因;私奔模型(runaway model)雄性对雌性的诱惑性。种群生态学种群生态学两种以不同歌声吸引伴侣的莺种群生态学螺蜗牛蛤种群生态学花园大蜗牛
豹类濒临绝境金钱豹雪豹黑豹种群生态学繁殖种的概念繁殖种的概念,集中在如下思想:物种延续并保持其遗传的完整性和独特性是因为不进行异种杂交。生物种概念和识别种概念都根据相互杂交定义种。本观点与实际的物种定义不同的是后者以明显的形态学性状为基础。
如热带蜗牛有两种,但是壳螺旋的方向不同。种群生态学生物种的概念生物种概念认为种是一组可以相互杂交的自然种群,它们与其他种群间具有繁殖隔离。该概念将没有明显形态学差别的种群分成分离的姊妹种,因为它们不能彼此交换基因。生物种由繁殖隔离机制来保持。种群生态学种群生态学异域性物种
形成与原来种由于地理隔离而进化形成新种,为异域性物种形成。异域性物种形成最易发生在边缘隔离、处在种分布区的极端边缘的小种群。小的非典型种群与极端环境条件的混合作用可产生迅速而广泛的遗传重组(遗传革命),从而导致物种形成。种群生态学邻域性物种
形成邻域性物种形成发生在分布区相邻,但分布区内不同地点环境(如气候)条件不同的种群。如在环布种(银鸥中的小黑背鸥和更小黑背鸥)中那样,可发现中间杂种,但包括的很大的距离使两种类型不能完全混合。AB C A: Macrpus giganteus; B: M. fuliginosus; C:M. rufus; D: Cicindela longilabrisD气候对三种袋鼠和一种虎甲分布的影响种群生态学同域性物种
形成同域性物种形成可能发生在没有地理隔离,但具有宿主选择差异、食物选择差异或生境选择差异的种群。同域性物种形成是否发生尚有争议,尽管已观察到植食性昆虫宿主选择的快速变化。植物可以通过多倍体进行同域性物种形成。
种群生态学是研究种群生物系统的规律的科学,具体地说,就是研究种群内部各成员之间,种群(或其成员)与其他生物种群之间,以及种群与周围环境非生物因素的相互作用规律。
种群生态学的重要内容之一是种群动态研究。种群动态研究也就是研究种群数量在时间上和空间上的变动规律及其变动原因。种群生态学 种群数量决定着该种群对生态系统作用的大小,对于有害动物或杂草、经济动物或植物、或是濒危物种,则相应反映着它们的危害程度、可利用程度以及濒危程度。
种群并不是简单的相加,种群作为一种新的生物系统应该有新质产生。种群生态学银杏 第一节 种群密度和分布
不同生物学分支学科对种群这一概念的理解存在着差异。
生态学上的种群概念强调了种群的分布和数量,并且强调了种群是一种生物系统,即种群不仅与环境相互关联,而且种群内部个体之间存在着遗传信息交换等种内关系。
种群生态学种群生态学种群密度很高种群密度一般种群生态学对于单体生物和种群如哺乳类,其种群大小就是一定区域内个体的数量,非常简单。对于构件生物,如植物和珊瑚,情况就较复杂。对于这些种群,“断片”(无性系分株)或枝条(构件)的数目比不同个体的数量更有意义,代表多度。种群大小种群生态学种群生态学种群的年龄结构是每一年龄阶段个体数目的比率,通常以年龄金字塔图来表示。既不增长也不下降的种群有稳定的年龄分布。增长型的种群有更多的年轻个体,而在下降型种群中年老的个体占优势。当种群经历离散的发育时期(如昆虫幼体的龄期)时,每一时期个体的数目(“时期结构”)可以对种群进行有效的描述。对于生长率无法预测的物种(如植物),根据大小分类可能更有意义。年龄和时期
结构 年龄锥体的三种基本类型
A增长型种群: 幼年组个体数多,老年组个体数少,种群的死亡率小于出生率,种群迅速增长。
B稳定型种群: 种群出生率大约与死亡率相当,种群稳。
C下降型种群: 幼年组个体数少,老年组个体数多,种群的死亡率大于出生率,种群种群数量趋向减少。繁殖后期繁殖期繁殖前期ABC种群生态学稳定年龄分布(stable age distribution)
年龄金字塔(age pyramid)
固定年龄分布(stationary age distribution)
植物体作为构件生物,有年龄结构,由年轻的、发育中的、正在起作用的和衰老的部分组成。企鹅种群生态学不同构件的植物种群生态学出生率就是新个体的产生,实际出生率就是一段时间内每个雌体实际的成功繁殖量。特定年龄出生率就是特定年龄组内雌体在单位时间内产生的后代数量。出生率死亡率是在一定时间段内死亡个体的数量除以该时间段内种群的平均大小。这是一个瞬时率,可用来估价整个种群的死亡率或特定年龄群的特定年龄死亡率,死亡的概率是死亡个体数除以在每一时间段开始时的个体数。死亡率种群生态学迁入率(immigration):进入当前种群领地的同种个体占总个体数的比率;
迁出率(emigration):离开当前种群领地的同种个体占总个体数的比率。
A型——凸型(convex curve)的存活曲线。 种群生态学B型——对角线型(diagonal straight 1ine)的存活曲线。 种群生态学C型——凹型(concave curve)的存活曲线。 种群生态学种群生态学存活率是死亡率的倒数。对于一个特定种群,存活率的数据通常以存活曲线的形式来表示;存活曲线表示的是在每一个生活期存活个体所占的比率的对数值。根据各种生活期死亡率的高低,特定年龄存活曲线一般有三种模式:后期死亡率最高(类型I),各期死亡率相等(类型II),早期死亡率最高(类型III)。存活率种群生态学树袋熊考拉种群大小随时间的变化可以按如下方法计算:t时间种群原来数量(Nt),加上新出生的个体数(B)和迁入个体数(I),减去死亡个体数(D)和迁出的个体数(E),就可得到t+1时间种群的数量(N t+1),这可用以下方程表示。
N t+1= Nt +B + I – D – E
在一组特定条件下,一个体具有最大的生殖潜力,称为内禀自然增长率r。这是种群在不受资源限制的情况下,于一定环境中可达到的理论最大值。种群生态学种群增长种群生态学种群生态学逻辑斯谛方程描述的是一个在有限资源空间中的简单种群的增长。在早期,资源丰富,死亡率最小,繁殖尽可能的快,种群内个体可达到内禀增长率。种群呈几何式增长,直到种群数量达到环境可持续支持的最大程度,这个最大数量称为环境容纳量(K)。当种群更加拥挤时,种群增长率减少到零,种群大小处于稳定状态。这可用逻辑斯谛方程来表示:
t时间种群大小变化率=内禀增长率×种群大小×密度制约因子
dN/dt = rN(1 - (N/K))
当种群达到环境容纳量,种内竞争变得更激烈时,密度制约因子(1 - (N/K))会接近零。该方程预测种群的增长随时间变化呈现出“S”形,如在真实种群中通常所观察的那样。密度制约性
种群增长:
逻辑斯谛方程种群的内禀增长率(intrinsic rate of increase):
自然增长率(rate of natural increase)是种群的实际增长率,用r来表示。它是指在单位时间内某一种群的增长百分比。
增长率=自然增长率+净迁移率=出生率-死亡率+迁入率-迁出率
内禀增长率或称为内在增长率(rm),即在无限制的环境条件下,种群的最大增长率。内禀增长率也称为瞬时增长率(instantaneous rate),或称为生物潜能(biotic potential)或生殖潜能(reproductive potential)。种群生态学苏格兰松和领鸽种群的指数增长曲线种群生态学 逻辑斯谛曲线可分为5个部分,即种群增长的5个时期:?
(1)开始期:潜伏期或缓慢期,初始种群刚刚适应环境。
(2)加速期:当初始种群适应环境后,个体加速繁殖,种群迅速增长,增长速度取决于初始种群的数量(N0)和瞬时增长率(r)。 ((3)转折期:种群增长达到最大后开始逐渐地减少。
(4)减速期:种群在这一时期的瞬时增长率迅速地下降。
((5)饱和期:种群密度接近或达到k值,不再继续增长。种群生态学 逻辑斯谛增长模型的意义:
(1)是许多两个相互作用种群增长模型的基础;
(2)是农、林、渔业等实践领域中,确定最大持续产量(maximum sustainable yield)的主要模型;
(3)模型中两个参数r和K已成为生物进化对策理论中的重要概念。生态密度(ecological density)单位面积(或体积)特定生态环境中的生物个体数。
根据密度调查方法的不同,密度可以分为绝对密度(absolute density)和相对密度(relative density)。种群生态学(一)绝对密度的常用调查方法
1.总数量调查法(total count)。又称直接统计法,是直接计数一定地方的所有个体,由此得到种群密度。E.g.:人口调查法(census technique)
2.取样调查法(sampling methods)。大多数绝对密度调查只是在几个地方计数种群的一小部分,由此估计种群整体的密度。
常用的取样调查法有三种:
①样方法(use of quadrate)
②标志重捕法(mark recapture methods):N=(M×n)/m
M是标志数,n是再捕个体数,m再捕中标记数
③去除取样法(removal sampling) :对于一个封闭的种群,对逐次捕获数/单位努力(作为y轴),逐次捕捉的累计数(作为x轴)作图,就可以得到一个回归线,到达 x轴的截距所表示的数量就是种群数量N。种群生态学(二)相对密度的调查方法
1.动物计数(counts of animals)
2.动物痕迹的计数(counts of animal signs)
3.单位努力捕获量(catch per unit effort)。
每一种生物的种群密度,都有一定的变化限度。
种群生态学白鹭计数时以其痕迹为依据 最大密度(maximum density)或称饱和密度(saturation density或upper asymptote level)是指特定环境所能容纳某种生物的最大个体数。当超过这一密度,种群数量将不再增长。
最小密度(minimum density或lower asymptote level)是指种群维持正常繁殖、弥补死亡个体所需要的最小个体数。如果低于最小密度,由于难以寻找配偶。繁殖意愿丧失、繁殖力下降,结果,种群就难以生存。
在最大密度和最小密度之间存在着一个最适密度(optimum density),当处于最适密度状态时,种群的增长最快。种群生态学处于饱和密度的森林种群生态学川金丝猴雪豹穿山甲犀鸟濒危动物性比率(sex ratio)是指种群中雄性与雌性个体数的比例。
大多数动物种群的性比接近1:1;
营社会生活的昆虫雄性多于雌性;
孤雌生殖的轮虫和枝角类等雌性多;
还有随环境条件的变化而性别改变的,水生动物如虾、黄鳝等。 种群生态学种群生态学轮虫河虾沼虾黄鳝种群生态学如果种群参数如出生率和死亡率随着种群密度的变化而变化,就可以说是密度制约的。如果出生率和死亡率不随着密度的变化而变化,则是非密度制约性的。如果没有迁移,除非出生率和死亡率是密度制约性的,否则种群会持续增长。
密度制约种群生态学种群生态学密度制约出生率和死亡率的平衡给出平衡密度,或环境容纳量 K。 (a)出生率和死亡率都是密度制约的;(b)非密度制约出生率;
(c)非密度制约死亡率;(d)同(a),但出生率和死亡率结合环境变化。种群生态学平衡种群密度当单位个体出生率正好平衡单位个体死亡率,种群密度既不增加也不减少时,此时的种群密度为平衡种群密度。平衡种群密度与环境容纳量K值相等。目前趋于平衡的斑马种群生态学密度制约有三种形式:过度补偿,补偿不足和准确补偿。如果是过度补偿,当初始密度上升时,存活个体数目将下降。如果密度制约是补偿不足,当初始密度上升时,存活个体数量上升。如果下降个体数与开始增加的个体数相等,则称密度制约为准确补偿。补偿和密度
制约表明的是密度制约性过程与种群对可利用资源的利用的对抗关系种群生态学大多数实际种群并不是在平衡密度停留很长时间,而是动态的不断的发生变化。由于周围环境的变化或生物因子的影响,种群可能会增长或收缩。增长种群和
收缩种群迁徙耕作种群生态学种群生态学种群生态学种群生态学混 沌数学上有关混沌的定义与俗语中的用法是截然不同的。一个混沌系统是受确定性作用控制的,而不是随机的。混沌的结果依赖于初始状态的精确值。由于完全精确是不可能达到的,因此对混沌系统不能进行有效的预测。一些分析认为,麻疹的爆发和一些昆虫种群可能表现出混沌的动态变化。种群生态学蟋蟀螳螂黑甲虫卵麻疹易于混沌的昆虫种群生态学季节消长:季节变动引起的自然种群数量变动,如北点地梅
种群爆发:具有不规则或周期性波动的生物都可能出现种群爆发,如害虫、赤潮等。
生态入侵(ecological invasion):由于人类有意识或无意识地把某种生物带入其适宜其栖息和繁衍的地区,该生物种群不断扩大,分布区逐步稳定地扩展,这种过程为生态入侵。水葫芦种群生态学蝗虫种群爆发种群生态学澳大利亚的侵入种群种群生态学种群的分布格局
组成种群的多个个体在其生活空间中的位置或布局,称为种群的内分布型(internal distribution pattern) :
(1)集群型(clumped)
(2)随机型(random)
(3)均匀型(uniform)树蕨,一种只分布于四川的植物集群型随机型均匀型种群生态学通常用泊松分布、负二项分布、奈曼分布等模型计算 第二节 种群的相互作用关系
一、集群
集群(aggregation或society, colony) 现象普遍存在于自然种群中。同一种生物的不同个体,或多或少都会在一定的时期内生活在一起,从而保证种群的生存和正常繁殖,因此集群是一种重要的适应性特征。
根据集群后群体持续时间的长短,可以把集群分为临时性(temporary)和永久性(permanent)两种类型。 种群生态学 生物产生集群的原因是复杂多样的,包括:
⑴对栖息地的食物、光照、温度、水等生态因子的共同需求;
⑵对昼夜天气或季节气候的共同反应。如过夜、迁徒、冬眠等群体;
⑶繁殖的结果;
⑷被动运送的结果;
⑸由于个体之间社会吸引力(social attraction)相互吸引的结果。
动物群体的形成可能是完全由环境因素所决定的,也可能是由社会吸引力(social attraction)所引起。种群生态学 集群的生态学意义:
(1)集群有利于提高捕食效率;
(2)集群可以共同防御敌害;
(3)集群有利于改变小生境;
(4)集群有利某些动物种类提高学习效率;
(5)集群能够促进繁殖。
拥挤效应(overcrowding effect)种群生态学种群生态学相互作用的
分类个体或物种间的相互作用可以相互作用的机制和影响为基础来分类。关键的种间相互作用是竞争、捕食、寄生和互利共生,而主要的种内相互作用是竞争、自相残杀和利他主义。偏利共生(commensualism):如寄居蟹与水螅虫,对后者有利,对前者无害;
偏害共生(amensualism):有毒物种和土壤真菌之间;
拟寄生(parasitoidism):通常指昆虫。种群生态学竞 争竞争是共同利用有限资源的个体间的相互作用,会降低竞争个体间的适合度。竞争即可在利用共同资源的物种间发生(种间竞争),也可在同种个体间发生(种内竞争)。个体或物种的生态位(它所处的环境,利用的资源和它发生的时间)是决定该个体或物种与其他个体或物种竞争程度的关键。大范围的生态位重叠一般导致激烈竞争。种群生态学当个体对资源的需要非常相似时,竞争会特别强烈。种内竞争是生态学的一种主要影响力,是扩散和领域现象的原因,并且是种群通过密度制约过程进行调节的主要原因。
如欧鳊,一种淡水鱼,幼鱼摄食小的浮游动物,成鱼以大型底栖无脊椎动物为食。种内竞争成鱼种群生态学种间竞争种间竞争发生在利用同样有限资源的两物种之间。极少种能够逃脱其他种与之竞争共同资源的影响。
如双核小草履虫和大草履虫在酵母介质中培养,结果前者占优胜,后者死亡等。种群生态学竞争有两种作用方式。在资源利用性竞争方式下,个体不直接相互作用,而是耗尽资源使供应不足。由于可利用资源不足而造成适合度下降。利用性竞争种群生态学在干扰性竞争方式下,个体直接相互作用,在一些动物种类中,最明显的通过打斗,也通过产生毒物(如植物异株克生)进行竞争。在这种相互作用中“败者”适合度下降,可能由于干扰(如受伤或死亡),也或许由于缺乏可用资源。干扰性竞争种群生态学他感作用:通常指一种植物通过向体外分泌代谢过程中的化学物质,对其他植物产生直接或间接的影响。许多高等植物能产生具有自毒(autotoxic)和抗生性质的有机物质,如碳水化合物中的萜烯、酒精、有机酸、醚、醛、酮等类物质。这些有机物质从各种途径释放出来,如根分泌直接进入土壤、从活或死组织淋洗出来、或挥发到空气中散布开来。这些物质能够抑制或促进其他物质的生长。
如:Encelia farinosa 一种菊科植物,叶子分泌的一种苯甲醛物质对相邻的番茄、玉米等物质有强烈的抑制作用;风信子、稠李、洋槐、丁香等的花,熏衣草、薄荷、月桂等的叶子的芳香物质对相邻植物伸长生长起抑制作用;榆属与栎树、白蜡树与栎树、白桦与松树、松树与云杉不能混种在一起;胡桃分泌物毒害苹果,对番茄和马铃薯也有影响;玉米对苹果有抑制作用;葱和菜豆、芜菁和番茄、番茄和黄瓜、冬黑麦和小麦之间都有抑制作用。
另一些植物,如洋葱和食用甜菜、马铃薯和菜豆、小麦和豌豆生长在一起却能彼此促进。正在猎食中的狮子领鸽(Streptopelia decaocto)的种群扩散过程自M.C.Molles,Jr,1999 非洲蜂(Apis melifera)的种群扩散自M.C.Molles,Jr,1999北美的枫树(Acer spp.)(左)和芹叶钩吻(Tsuga canadensis)(右)的种群扩散自M.C.Molles,Jr,1999种群生态学许多动物种(包括昆虫、鸟类和哺乳动物)的个体或群体为争夺空间而竞争。保持领域边界的个体间有积极的相互干扰行为。领域使其所有者受益,为对抗入侵者,保护领域所付出的代价,被增加的食物供应、提高的交配成功率和降低的被捕食的危险所带来的价值超过。领 域 性社会等级(social hierarchy):动物种群中的支配和从属关系。保卫“我”的领地种群生态学固着生长的生物,包括植物,不能通过运动逃避竞争,因此竞争中的失败者死去在同样年龄大小的植物群中,这种竞争结果使较少量的较大个体存活下来。这一过程叫做“自疏”。自疏导致密度与植物个体大小之间的关系,这在双对数作图时呈现典型的-3/2斜率。这种关系叫做Yoda-3/2自疏法则。自 疏种群生态学菊花的自疏种群生态学如果两个物种在稳定环境中竞争,则有两种可能的结果:(i)一种被排除,或(ii)两种共存。竞争排斥原理陈述,共存只能发生在两物种生态位分化的稳定环境中。竞争排斥种群生态学In the absence of a competitor, a species may expand its niche. Examples of such competitive release include ground doves in New Guinea and gerbils in Israel.Competitive release种群生态学在缺乏竞争者时,物种会扩张其实际生态位。这种竞争释放的例子包括新几内亚岛上的地鸽子和以色列的沙鼠。竞争释放种群生态学当明确的生态位契约受到竞争影响时,为了适应新的资源谱,个体形态会相应变化。这种性状替换发现在收获蚁Veromessor Pergandei 和达尔文雀Geospiza fortis和G. fuliginosa。性状替换种群生态学环境是许多不同质量、不同资源水平的生境的拼缀。一些斑块的发生是暂时的和不可预测的。在时间和空间上,环境是异质性的。这意味着竞争“战斗”在环境变化和竞争平衡变化之前不会完成。因此,在现实世界中共存的种类,其共存可能是因为环境的异质性,而不是因为生态位分化。空间和时间
异质性种群生态学如果捕食者进攻两种猎物,那么每一种猎物,会通过增加当地捕食者种群的数量,相反地影响另一种。因此,两个猎物种群之间的相互作用看上去似乎它们确实是在竞争,然而它们或许利用完全不同的资源。这一现象叫做表观竞争。表观竞争种群生态学五、捕食种群生态学捕食可定义为摄取其他个体(猎物)的全部或部分为食。这一广泛的定义包括
(i)典型的捕食者,在袭击猎物后迅速杀死而食之;
(ii)草食者,只消费对象个体的一部分;(iii)寄生者,与单一对象个体(寄生)有密切关系,通常生活在寄主的组织中。捕食的定义等待猎物的老虎种群生态学捕食者可以划分为(i)消费植物组织的草食者,(ii)摄食动物组织的肉食者和(iii)既摄食植物组织也摄食动物组织的杂食者。动物、植物食性的差异需要不同的生理和行为适应,从而导致肉食者与草食者世系之间反复的进行分歧。肉食者和
草食者种群生态学捕食者随其摄取猎物的数量多少而变化,某些捕食者是特化种,而另一些是泛化种。一般来说,寄生者比典型捕食者更为特化、而草食者比肉食者更为特化。泛化种和
特化种种群生态学种群生态学捕食者和寄生者是否能够调节其猎物种群的大小呢?这一问题并不像看上去的那么简单。有两种主要观点:(i)任一捕食者的作用,只占猎物种总死亡率的很小一部分,因此去除捕食者对猎物种仅有微弱效果;(ii)捕食者只是杀死了对象种中即将死亡的个体,所以最终对猎物种群大小没有影响。然而,在一些例子中确有明显证据表明捕食者对猎物数量有重要影响。捕食者对猎物
种群大小的
影响种群生态学种群生态学捕食者-猎物模型行为多度捕食者猎物时间N种群生态学捕食者和猎物的种群动态分为4个时期:a猎物增加,捕食者减少;b猎物和捕食者都增加;c猎物减少,捕食者继续增加;d猎物和捕食者都减少。种群生态学猎物收益率种群生态学猎物转换依据猎物种数量的多少,捕食者有时会转换其选择而捕食某一特定的猎物种。这时捕食者主要捕食优势种猎物而大大忽略其他猎物。猎物密度影响
——功能反应一般认为在高猎物密度下,捕食者的摄食率会增加,然后随猎物饱和达到最大速度。这种关系称为功能反应,可能采用多种模式,传统上分为三种类型:功能反应I、II和III。种群生态学Ⅰ随猎物密度的增加,捕食者的摄食以固定比例增加;Ⅱ捕食者在摄食猎物时可能达到饱足,摄食率降低,直到完全饱足,摄食率不再随猎物密度的变化而变化;Ⅲ捕食者在猎物密度低时由于低狩猎效率而反应低,以后摄食率又因饱足水平而减少。种群生态学搜寻和处理为得到食物,捕食者必须首先搜寻猎物,然后处理(抓住、加工和吃掉猎物)。可以认为捕食者食谱的宽度是由泛化种对策与特化种对策之间的权衡决定的。泛化种对策捕食者寻找多种猎物(相对容易),特化种对策捕食者寻找一类猎物,非常有效的处理它。最佳觅食理论假定进化会最优化动物行为以使其获得的能量效率最大,从而做出捕食者如何权衡搜寻与处理的预测。种群生态学狩猎者 种群生态学种群生态学种群生态学疾藜草─是兇惡植
物,身上插滿一根根
堅硬又尖銳的針刺。
任何動物只要一不小
心惹上它們就會疼痛
不已。种群生态学寄生物的
多样性寄生物是摄取其他生物的组织、与其寄主紧密相联而生的捕食者的子群。寄生物可以分为两大类(i)微寄生物,在寄主体内或表面繁殖,(ii)大寄生物,在寄主体内或表面生长,但不繁殖。主要的微寄生物有病毒、细菌、真菌和原生动物。寄生蠕虫和昆虫是重要的大寄生物。一大类昆虫大寄生物是拟寄生生物,它们在昆虫寄主身上或体内产卵,通常导致寄主死亡。鸡肠中的蛔虫种群生态学种群生态学种群生态学SARS病毒种群生态学传播方式寄生物可以横向传播(在种群个体之间),或在少数情况下,纵向传播(从母体到后代)。横向传播或直接或间接,由传病媒介(如蚊子)或中间寄主做中介。种群生态学寄主对疾病的反应脊椎动物被微寄生物感染后产生强烈的免疫反应。这种反应主要有两种成分:(i)细胞免疫反应,特定细胞直接攻击病原体细胞,和(ii)B-细胞免疫反应,使抗体量上升。初次被病原体感染后,免疫记忆对未来同样的侵袭产生快速反应,导致免疫。无脊椎动物和植物也可以抑制感染,但方式较简单,通常较少特化机制。免疫造成的潜在寄主减少,使微寄生物的繁荣-和-破产对策加强.种群生态学疾病的传播途径种群生态学种群生态学社会性寄生物:“社会性寄生物”与其寄主之间有一种完全不同的关系。社会性寄生物不通过摄食寄主组织获益,而是通过强迫寄主提供食物或其他利益而获利。这种关系发现在杜鹃,它们将蛋下在别种鸟的巢中,让这种鸟饲育其幼鸟。这种关系还发现在一些蚂蚁种类,它们迫使其他种工蚁给它们的幼体食品或其他利益。
种群生态学工蚁种群生态学种群生态学种群生态学蜜蜂与花的协同进化种群生态学寄主——
寄生物进化寄生物与其寄主间紧密的关联经常导致二者在进化上的相互作用,或协同进化。协同进化使寄主的防御机制提高,而给寄生物克服这些防御的通路——这叫做“军备竞赛”。但是,寄主与寄生物之间的协同进化并不都是提高性的。例如,协同进化会降低寄生物的毒性。 大熊猫体内存有纤毛虫种群生态学寄生物种群的异质性一些疾病是稳定的,而另一些进化了通过变异“逃离”寄主防御的手段。人类“孩童时期”的疾病——麻疹、腮腺炎、水痘等大部分局限于孩子,因为它们的抗原稳定。与此相对,寄生物如流感病毒、鼻病毒(普通感冒原)和沙门氏菌(Salmonella)具有可经常加入的很多变异型。寄主因此要经常与变化的寄生物军队的进攻而进行战斗。种群生态学种群生态学七、互利共生( Mutualism)互利共生和
共生现象互利共生是不同种两个体间一种正的互惠关系,可增加双方的适合度。互利共生可以是共生性的,生物体以一种紧密的物理关系生活在一起。种群生态学地 衣种群生态学蝴蝶传粉种群生态学种群生态学传 粉异型杂交植物需要将其花粉转移到同种植物的柱头上,并接受同种植物的花粉。一些植物种类靠风来传播花粉,如果植物生长在广大的植物种类单一的地方如草地和松林,这种传播方式会很有效。但是大多数开花的双子叶植物,依靠昆虫、鸟、蝙蝠或小型哺乳动物来传播花粉,作为交换,这些动物或以花蜜或以花粉本身为食。种群生态学传 粉种群生态学两种蜂鸟及其特化对应的花的形态种群生态学种子扩散大型种子无法有效的被风传布,除非靠水传布(如椰树的种子),或依靠动物来散布。啮齿动物、蝙蝠、鸟和蚂蚁都是重要的种子扩散者。
食水果者(frugivore)来自澳洲的老鼠种群生态学浦公英种群生态学互利共生和
进化互利共生可能起源于寄生物-寄主和捕食者-猎物之间的关系,或起源于没有协作或相互利益的紧密共栖物种之间。然后双方的进化变化(协同进化)导致双方从这种关系中获益,虽然也可能共生“恶化”,成为使一方受益、另一方受非平衡的剥削的寄生关系。互利共生的相互作用在多细胞动物进化过程的一些关键阶段中起着中心作用。高等生物的许多细胞器官,包括线粒体和叶绿体(都来自自由生活的原核生物,含有环状DNA基因组和其他细胞特征),被认为来自共生性细胞。互利共生的例子:人与家畜(不能独立),丝暇虎鱼利用鼓虾的巢穴作隐蔽场所,鼓虾(不能独立)是盲的,离开洞穴时,以细长的触角保持与鱼的接触,代替眼睛起警报作用。后者如花和蜜蜂,反刍动物与瘤胃内的细菌和原生动物,高等植物与真菌(菌根),白蚁和肠道内的鞭毛虫等。
红豆杉大量繁殖的竹子种群生态学 迁移:生物也可以通过移动到别处来躲避当地恶劣的环境。迁徙是方向性运动,如燕子从欧洲到非洲的秋季飞行。相反,扩散是离开出生或繁殖地的非方向性运动。迁移有三种,依据生物个体做的是(i)反复的往返旅行,(ii)单次往返旅行(死之前回道出生地进行繁殖),或(iii)单程旅行(一个世代向一个方向运动,下一个世代返回来)。
迁徙中的鸟种群生态学种群生态学种群生态学复杂的生活
周期许多植物、真菌和动物具有复杂的生活周期,在生活周期中,或者个体采用根本不同的形态学性状(如毛虫/蝴蝶,蝌蚪/青蛙)或者以一种可预测的方式采取彼此根本不同的世代(如植物的单倍体/双倍体世代交替,锈菌和蚜虫的有性/无性世代交替)。为什么要采用这些复杂的生活史对策呢?在一些种类、不同的时期,或者主要进行生长,或者主要进行扩散,而在其他种类,不同的时期是为了最有效地利用不同的生境。种群生态学衰 老为什么生物个体变老后身体恶化,繁殖力和精力降低?对这一问题有两个水平的答案。在机制水平上,由于毒物和自然辐射的影响,使细胞机器崩溃,从而引起衰老。但是,这不能是全部的原因——衰老的发生随种类不同变化很大。这说明进化影响或决定着衰老。衰老的进化模型认为,或者(i)一个影响较老个体的有害突变,比影响年轻个体的有害突变,受到更弱的选择对抗,或者(ii)对早期繁殖有益的基因在生命晚期有恶性影响。
又称为突变积累(mutation-accumulation),早期表达的基因会显著降低个体的繁殖输出;拮抗性多效(antagnistic pleiotropy),如马鹿的一个与提早繁殖和高繁殖力相关的基因也与低存活力有关,而黑尾果蝇对提高的存活力的选择使早期繁殖力降低。默契的天鹅种群生态学合作的蚂蚁种群生态学加里福尼亚鸟群蜂王种群生态学生态学中的性性性是植物和动物中广泛分布的现象,性有一系列生态后果。性在进化上的重要性是由于它产生遗传变异,它在生态学上的重要性则是因为不同性别的生物其行为不同。两性之间的区别是由于它们对后代发育的能量投资不对称性而产生的。(很多雄性仅提供精子或花粉,雌性却要提供大的卵,子宫胚珠和乳汁等)种群生态学近交的代价当亲缘相近的个体进行近亲繁殖时,“近交”就出现了。近交引起(i)近交衰退(它使后代的适合度降低)和(ii)增加遗传纯合性(它降低了对于环境变化产生进化适应的可能性,而增加了灭绝的可能性)。(有害隐性等位基因的纯合频率的增加在亲缘关系近的种群中更容易出现,是近交衰退的原因。)许多物种具有防止或减少近交的机制。笼养动物和野生动物种群生态学这是全球发现的第一只饲养中雌雄同体的蝴蝶。这只大凤蝶最近从新加坡圣陶沙的蝴蝶园内破蛹而出。种群生态学交配体制动物,特别是脊椎动物,其交配的社会结构是重要的生态特征。一个雄体和一个雌体形成配偶,或者仅在繁殖季节,或者直到其中有一个死亡,这是单配制(monogyny)(鸟类)。当一个雄体与多个雌体交配,就出现了一雄多雌制(polygyny)(哺乳类);当出现相反情况(很少见),即一个雌体具有多个雄体的交配群,就叫做一雌多雄制(polyandry)(北极冻原的鸻科鸟和最严酷的喜马拉雅地区的人类社会)。种群生态学种群生态学性 比性比是种群中雄体对雌体的比率,通常接近于1雄:1雌。但是,不等的消耗(条蜂,雌性比雄性重58%,理论值性比为1.58:1,实测值为1.63:1)、拥挤的配偶竞争(雌体间的局域资源竞争紧张,产出的雄体离家出走)也可能使性比偏离。种群生态学在孔雀的种群中,雄性个体比雌性多得多种群生态学性 选 择性选择导致形成像孔雀之尾、雄苇莺动听的歌唱、雄鹿的叉角等特征。交配竞争中生殖成效的不同导致性选择。
有性内选择(打斗)和性间选择(偏爱异性的某个独特特征。让步赛理论(handicap theory)即好性状有好基因;私奔模型(runaway model)雄性对雌性的诱惑性。种群生态学种群生态学两种以不同歌声吸引伴侣的莺种群生态学螺蜗牛蛤种群生态学花园大蜗牛
豹类濒临绝境金钱豹雪豹黑豹种群生态学繁殖种的概念繁殖种的概念,集中在如下思想:物种延续并保持其遗传的完整性和独特性是因为不进行异种杂交。生物种概念和识别种概念都根据相互杂交定义种。本观点与实际的物种定义不同的是后者以明显的形态学性状为基础。
如热带蜗牛有两种,但是壳螺旋的方向不同。种群生态学生物种的概念生物种概念认为种是一组可以相互杂交的自然种群,它们与其他种群间具有繁殖隔离。该概念将没有明显形态学差别的种群分成分离的姊妹种,因为它们不能彼此交换基因。生物种由繁殖隔离机制来保持。种群生态学种群生态学异域性物种
形成与原来种由于地理隔离而进化形成新种,为异域性物种形成。异域性物种形成最易发生在边缘隔离、处在种分布区的极端边缘的小种群。小的非典型种群与极端环境条件的混合作用可产生迅速而广泛的遗传重组(遗传革命),从而导致物种形成。种群生态学邻域性物种
形成邻域性物种形成发生在分布区相邻,但分布区内不同地点环境(如气候)条件不同的种群。如在环布种(银鸥中的小黑背鸥和更小黑背鸥)中那样,可发现中间杂种,但包括的很大的距离使两种类型不能完全混合。AB C A: Macrpus giganteus; B: M. fuliginosus; C:M. rufus; D: Cicindela longilabrisD气候对三种袋鼠和一种虎甲分布的影响种群生态学同域性物种
形成同域性物种形成可能发生在没有地理隔离,但具有宿主选择差异、食物选择差异或生境选择差异的种群。同域性物种形成是否发生尚有争议,尽管已观察到植食性昆虫宿主选择的快速变化。植物可以通过多倍体进行同域性物种形成。
同课章节目录
- 第一章 植物生命活动的调节
- 第一节 植物激素调节
- 第二节 其他调节
- 第二章 动物生命活动的调节
- 第一节 内环境与稳态
- 第二节 神经系统的结构与功能
- 第三节 高等动物的内分泌系统与体液调节
- 第三章 免疫系统与免疫功能
- 第一节 人体对抗病原体感染的非特异性防卫
- 第二节 特异性反应(免疫应答)
- 第三节 免疫系统的功能异常
- 第四章 种群
- 第一节 种群的特征
- 第二节 种群的增长方式
- 第三节 种群的数量波动及调节
- 第五章 群落
- 第一节 群落的物种组成和优势种
- 第二节 植物的生长型和群落结构
- 第三节 物种在群落中的生态位
- 第四节 群落的主要类型
- 第五节 群落演替
- 第六章 生态系统
- 第一节 生态系统的营养结构
- 第二节 生态系统中的生产量和生物量
- 第三节 能量流动和物质循环
- 第四节 生态系统的稳态及其调节
- 第七章 人类与环境
- 第一节 生物圈
- 第二节 全球人口动态
- 第三节 人类对全球环境的影响