高中生物课件浙教版必修2第3章第3节:《遗传信息的传递》
文档属性
| 名称 | 高中生物课件浙教版必修2第3章第3节:《遗传信息的传递》 |

|
|
| 格式 | rar | ||
| 文件大小 | 476.8KB | ||
| 资源类型 | 教案 | ||
| 版本资源 | 苏教版 | ||
| 科目 | 生物学 | ||
| 更新时间 | 2010-11-16 00:00:00 | ||
图片预览









文档简介
课件51张PPT。
遗传信息的传递
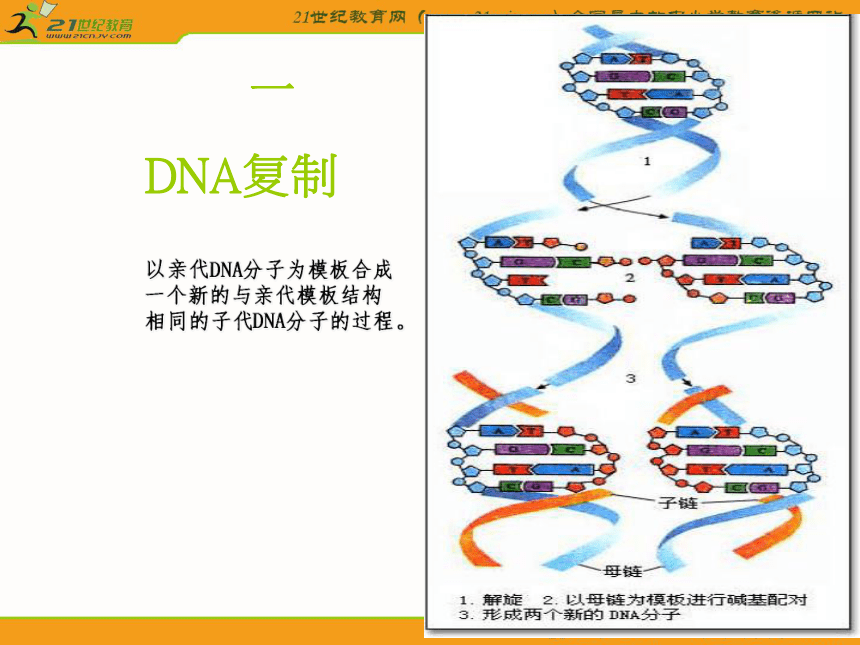
一、DNA复制
二、DNA的转录
三、蛋白质的生物合成
四、基因表达调控
五、基因与性状 遗传信息的传递
主要内容遗传信息的传递概叙遗传信息的传递(中心法则)包括以下过程:DNA复制、DNA转录(逆转录、RNA复制)、蛋白质合成一
DNA复制
以亲代DNA分子为模板合成一个新的与亲代模板结构相同的子代DNA分子的过程。2.半不连续合成
DNA的双螺旋结构中的两条链是反向平行的,当复制开始解链时,亲代DNA分子中一条母链的方向为5′~3′,另一条母链的方向为3′~5′。DNA聚合酶只能催化5′~3′合成方向。在以3′~5′方向的母链为模板时,复制合成出一条5′~3′方向的前导链,前导链的前进方向与复制叉的行进方向一致,前导链的合成是连续进行的。而另一条母链仍以3′~5′方向作为模板,复制合成一条5′~3′方向的随从链,因此随从链合成方向是与复制叉的行进方向相反的。随从链的合成是不连续进行的,先合成许多片段,即冈崎片段。最后各冈崎片段再连接成为一条长链。由于前导链的合成连续进行的,而随从链的合成是不连续进行的,所以从总体上看DNA的复制是半不连续复制。(二)、 DNA复制所需的酶及蛋白质
1.DNA聚合酶
DNA复制时,以DNA为模板,以A、T、C、G为底物,按照碱基互补配对原则,沿5’→3’方向合成新的DNA双链。
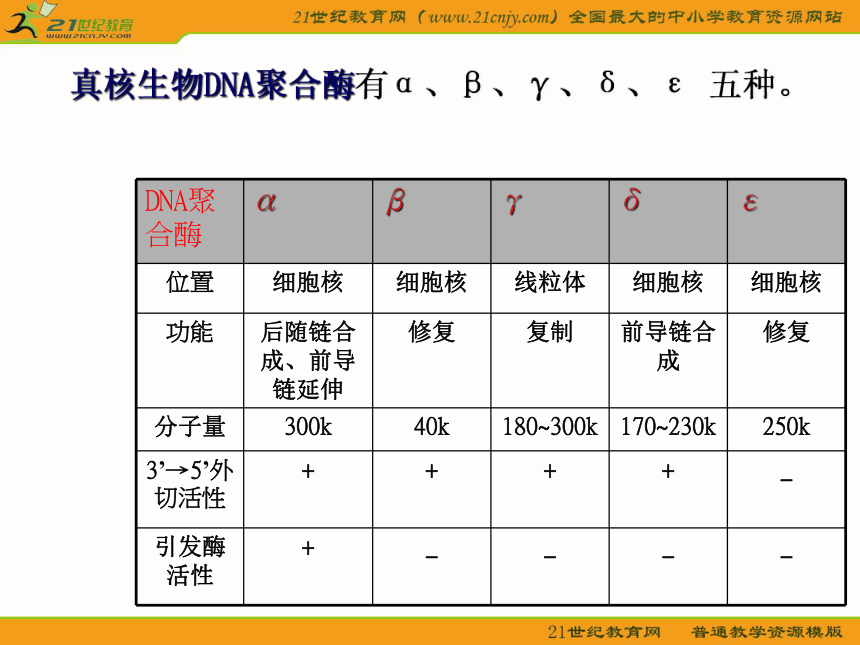
原核生物DNA聚合酶包括:DNA聚合酶Ⅰ:具有三种功能—即5’→3’ 聚合酶活性、 3’→5’外切校正活性、 5’→3’外切酶活性; DNA聚合酶Ⅱ;DNA聚合酶Ⅲ: 细菌DNA复制的主要聚合酶,具有5’→3’ 聚合酶活性、 3’→5’外切校正活性。真核生物DNA聚合酶有α、β、γ、δ、ε 五种。2.引发酶
起始DNA复制。真核生物DNA聚合酶Ⅰ具有引发酶活性;原核生物由dnaG基因编码。
3 .DNA连接酶
将后随链中的各冈崎片段连接成一条完整的互补链。
4、拓扑异构酶
消除、维持、形成DNA的超螺旋结构,分Ⅰ、Ⅱ两种。 5、解链酶
解开DNA分子的互补双链作为复制的模板。
6、单链结合蛋白
结合解开的单链DNA,保护其不被水解和恢复DNA双链结构,又称为双螺旋反稳定蛋白。(三)、 DNA复制的过程
1.DNA复制起始:蛋白质-DNA复合物形成,复制叉及引发体产生。
2、DNA复制延伸:复制叉前移、前导链及后随链延伸。
3、DNA复制的终止:RNA引物切除、缺口补齐、冈崎片段连接。(四)、原核生物及真核生物DNA复制的比较
1.共同点:
(1)都具有半保留和半不连续特征;(2)都需要特定的酶和蛋白质;(3)复制过程都包括起始、延伸和终止三个阶段。
2、不同点:
(1)复制过程所需要的酶和蛋白因子不同;(2)原核生物DNA为环状分子,采用θ型复制和滚环式复制;真核生物通过端粒酶识别和结合端粒,维持线性DNA分子的恒定长度;(3)原核生物只有1个复制起点,复制速度快、冈崎片段长,能连续复制;真核生物有多个复制起点,复制速度慢、冈崎片段短,不能连续复制。二、DNA的转录 (Transcription)以DNA为模板,A、U、C、G为底物,按照碱基互补配对原则从5’→3’方向合成RNA的过程,称转录。2、真核生物的RNA聚合酶2.mRNA前体的加工
(1)5′末端帽子的生成
过程:一个m7G连接到mRNA的5‘端形成(脱Pi、甲基化),真核生物有0、1、2三种类型帽子结构。
功能:① 维持mRNA的一级结构,避免磷酸酶和核酶的攻击; ② 提供核糖体的识别部位。
(2)3′末端多聚A尾的生成
通过识别加尾信号AATAAA,在其下游20bp处加上100-200个A,形成polyA尾巴;
功能:①增加mRNA的稳定性;②方便mRNA从细胞核进入细胞质;③增强翻译效率;(3)剪接作用
在核酸内切酶作用下剪切掉内含子,然后在连接酶作用下,连接各部分外显子。
(4) RNA编辑
在转录产物中插入、删除或取代一些核苷酸残基,生成具有正确翻译功能的模板,此即所谓 RNA的编辑作用。 编辑过程由一个或多个小分子的“指导RNA”提供mRNA的编辑信息,并作为模板指导其进行编辑,在编辑体的帮助下进行编辑。(5)甲基化修饰
原核生物mRNA分子中不含有稀有碱基,但真核生物的mRNA中则含有甲基化核苷酸,除了在hnRNA的5′端帽子结构中含有2-3个甲基化核苷外;在分子内部还会有l-2个m6A存在于非编码区。在序列中, m6A总是位于胞苷之后,形成了…NC m6A N序列。 m6A的生成是在hnRNA的剪接作用之前发生的。
真核生物的CpG岛的C容易形成甲基化,常常突变成T。在基因的末端通常存在一些富含双核苷酸“CG”的区域,称为“CpG岛”(CpG island)。在人类基因组内,存在有近3万个CpG岛;在大多数染色体上,平均每100万碱基含有5~15个CpG岛,其中有1.8万多个CpG岛的GC含量为60%~70%。通常,这些CpG岛不仅是基因的一种标志,而且还参与基因表达的调控和影响染色质的结构。Epigenetics3.tRNA的转录后加工
①在核酸内切酶 RNaseP作用下,从5′末端切除多余的核苷酸;
②在核酸外切酶 RNaseD作用下,从3′末端切除多余的核苷酸;
③核苷酸转移酶催化, 3′末端加CCA-OH,为tRNA加I特有反应;
④核酸内切酶催化进行剪切反应,剪掉内含子,由连接酶连接外显子部分;
⑤ 化学修饰作用,如甲基化、脱氨基、还原反应。
4. rRNA的转录后加工
原核生物有 16S、23S及 5S三种 rRNA,这三种rRNA均存在于30S的rRNA前体中。转录作用完成后,在RNaseⅢ催化下,将rRNA前体切开产生16S、25S及 5S rRNA的中间前体。进一步在核酸酶的作用下,切去部分间隔序列,产生成熟的 16S、23S及5S rRNA,还有成熟的 tRNA。并对16S rRNA进行甲基化修饰,生成稀有碱基。与4S rRNA加I变化不大。
真核生物的核蛋白体中有18S、5.8S及5S rRNA。 5SrRNA自己独立成体系,在成熟过程中加工甚少,不进行修饰和剪切。 45S rRNA前体中包含有 18S、5.8S及 28SrRNA。在加工过程中,分子广泛地进行甲基化修饰,主要是在28S及18S中。甲基化作用多发生于核糖上,较少在碱基上。随后45 S rRNA前体经核酸酶顺序剪切下生成18S、5.8S、28S rRNA。
三、蛋白质的合成-翻译 (Translation) 基因的遗传信息在转录过程中从DNA转移到mRNA,再由mRNA将这种遗传信息表达为蛋白质中氨基酸顺序的过程叫做翻译,即蛋白质的合成。
(一)、 蛋白质合成体系
1、mRNA提供遗传密码
在mRNA中,每3个相互邻接的核苷酸,其特定排列顺序,在蛋白质生物合成中可被体现为某种氨基酸或合成的终止信号者称为密码子,统称遗传密码。它具有连续性、简并性、通用性。UAA、UAG、UGA为肽链的终止信号,不代表任何氨基酸,称为终止密码子;密码子AUG不仅代表一定氨基酸,而且位于mRNA启始部分,称起始密码子。编码同一氨基酸的称同义密码子。起编码氨基酸作用的称有义密码子。
遗传密码及相应的氨基酸2、tRNA转运氨基酸
tRNA上有由3个核苷酸组成的反密码子,与mRNA上的密码子按碱基互补配对原则结合, tRNA与对应氨基酸结合成为氨基酰tRNA,氨基酰tRNA才能准确的在mRNA上对号入座。
3、核糖(蛋白)体提供装配场所
核蛋白体相当于装配体,当核蛋白体在mRNA上每向前移动一个密码子的位置,肽链上即增加一个新的氨基酸,直至终止信号。
4、氨基酸为合成原料
20种氨基酸(二)、 蛋白质合成过程
1.启动阶段
在蛋白质生物合成的启动阶段,核蛋白体的大、小亚基,mRNA与一种具有启动作用的氨基酸tRNA共同构成启动复合体。
2.肽链延长阶段
根据 mRNA上密码子的要求,新的氨基酸不断相应的被特异的tRNA运至核蛋白体受位,形成肽键。
3.终止阶段
当多肽链合成已完成,并且“受位”上已出现终止信号,此后即转入终止阶段。终止阶段包括已合成完毕的肽链被水解释放,以及核蛋白体与tRNA从mRNA上脱落的过程。(三)、 翻译后加工
1.一级产物的修饰:(1)去除N-甲酰基或N-蛋氨酸;(2)个别氨基酸的修饰;(3)水解修饰;(4)切除部分肽链(连接肽、信号肽)。
2.高级结构的修饰:肽链释放后可自行根据其一级结构的特征折叠、盘曲成高级结构。此外,高级结构的修饰还包括(1)折叠、(2)亚基聚合、(3)辅基连接等行为。
3.蛋白质合成的靶向输送:蛋白质合成后,定向地到达其执行功能的目标地点,称为靶向输送;(2)切除。四、基因表达的调控(一)、 基因及其结构
1.基因概念的发展
(1)1866年孟德尔在《植物杂交试验》中提出的遗传因子概念,是基因雏形名词。 (2)1909年丹麦遗传学家约翰逊在《精密遗传学原理》中提出“基因”概念来替代孟德尔假定的“遗传因子”。
(3) 1926年摩尔根的巨著《基因论》出版,提出基因以直线形式排列,决定特定性状,能发生突变和交换,它不仅是决定性状的功能单位,而且是一个突变单位和交换单位。
(4) 1957年法国遗传学家本滋尔提出顺反子学说,认为基因是DNA分子上一段核苷酸顺序,负责着遗传信息传递。2.基因的几个相关概念
(1)基因:可转录一条完整的RNA分子或编码一条多肽链的一段DNA序列;产物具有一定的生物学功能;
(2)假基因(pseudo gene):同已知的基因相似,但位于不同位点,因缺失或突变而不能转录或翻译,是没有功能的基因。
(3)等位基因(allele):位于同源染色体上,位点相同,控制着同一性状的基因。
(4)结构基因(structural gene):可编码RNA或蛋白质的一段DNA序列。
(5)调控基因(regulator gene):其产物参与调控其他结构基因表达的基因。
(6)重叠基因{overlapping gene}:同一段DNA的编码顺序,由于阅读框架的不同或终止早晚的不同,同时编码两个或两个以上多肽链的基因。(7)隔裂基因(split gene):一个结构基因内部为一个或更多的不翻译的编码顺序,如内含子(intron)所隔裂的现象。
(8)跳跃基因(jumping gene):可作为插入因子和转座因子移动的DNA序列,也称转座因子。
(9)主基因(major gene):对于性状的作用比较明显,容易从杂种分离世代鉴别开来。
(10)修饰基因(modifying gene):一组效果微小的基因能增强或削弱主基因对表型的作用,这类微效基因在遗传学上称为修饰基因。
(11)管(持)家基因:某些基因是维持细胞基本代谢所必须的基因,其在一个生物个体的几乎所有细胞中持续表达,称为~;
(12)奢侈基因:有些基因在一些分化细胞中活动,是细胞分化、生物发育的基础,称为~。 3.基因的结构5‘侧翼区(5‘ –Flanking Region) :基因5‘端部到转录起始点间的部分;
5‘UTR: 5‘非翻译区(Un-translated Region);
启动子:指RNA聚合酶结合并起动转录的DNA序列,通常包括TATA框等结构;
转录起始点
转录终止点
起始密码子(Start Codon):通常为ATG;
终止密码子(Stop Codon):通常为TAA/G;
Poly(A)信号:决定加尾(转录终止)位置;
外显子(Exon):出现在成熟RNA中的有效DNA区段;
内含子(Intron):基因内部的部分序列并不出现在成熟mRNA中,这些间隔序列称为内含子, 符合“GT-AG”规则。真核生物基因的普通结构(二)、基因表达的概叙
1、概念
基因表达就是基因转录及翻译的过程。在一定调节机制控制下,大多数基因经历基因激活、转录及翻译等过程,产生具有特异生物学功能的蛋白质分子,但并非所有基因表达过程都产生蛋白质, tRNA、rRNA编码基因转录合成RNA的过程也属于基因表达。
2、基因表达的时间性及空间性
基因表达的时间、空间特异性由特异基因的启动子和增强子与调节蛋白相互作用决定。(1)时间特异性:按功能需要,某一特定基因的表达严格按特定的时间顺序发生,这是基因表达的时间特异性。 多细胞生物基因表达的时间特异性又称阶段特异性。(2)空间特异性:在个体生长全过程,某种基因产物在个体按不同组织空间顺序出现,这就是基因表达的空间特异性。又称细胞特异性或组织特异性。4、基因表达的复杂性表现为(1)多级调控:染色体和基因的结构活化、转录起始、转录后加工及转运、mRNA降解、翻译及翻译后加工及蛋白质降解等均为基因表达调控的控制点。(2)多要素综合作用:DNA序列、调节蛋白和RNA聚合酶等的结合作用。
5、基因表达的生物学意义:主要体现在适应环境,维持生长、增殖、个体发育和分化等方面。(三)、原核生物基因表达调控
1、操纵子学说
--原核生物的基因表达可在 DNA水平、转录水平和翻译水平三个层次进行调控,其中转录水平调控是基因表达调控的主要环节;
--操纵子是原核生物转录水平调控的主要方式,操纵子由调节基因R、操纵基因O和结构基因S组成;
--操纵子通过调节基因编码的调节蛋白开启或关闭操纵基因,调控结构基因的表达;
--如果调节蛋白关闭基因表达活性,称为负调控;反之,调节蛋白开启基因表达活性,称为正调控;
--一些小分子物质可作用于调节蛋白,开启转录活性的为诱导物,反之为辅阻遏物、超阻遏物。A转
乙
酰
酶?-
半
乳
糖
苷
透过酶ZY2、操纵子结构和功能
--乳糖操纵子模型;
--乳糖操纵子的结构;P’ R--乳糖操纵子的功能(负调控机制)3、操纵子的其他调控形式
(1)色氨酸操纵子;
(2)半乳糖操纵子(gal operon)
(3)阿拉伯糖操纵子(ara operon)
4、基因表达的时空调控
噬菌体SPO1侵染枯草杆菌:侵染后,早期基因立即转录;4-5m后,转为中期基因表达;8-12m后转为晚期基因表达。
5、DNA序列重排的调控
DNA序列的重排调控基因的表达,如鼠伤寒沙门氏菌的相转变机制(包括P的DNA倒位)。(四)、真核生物基因表达调控
1、真核基因组结构特点:基因组(Genome)是指一特定生物体的整套(单倍体)遗传物质的总和 。
(1)真核基因组结构庞大:哺乳类动物基因组DNA长达10 9个bp。
(2)单顺反子:真核基因转录产物为单顺反子,即一个编码基因转录生成一个 mRNA分子,经翻译生成一条多肽链。顺反子表示一个起作用的单位,其所包括的一段DNA与一个多肽链的合成相对应,是基因的基本功能和转录单位,一个基因可有几个顺反子,一个顺反子产生一条mRNA。
(3)重复序列:在原核、真核DNA中都有重复出现的核苷酸序列,但在真核更普遍。根据重复频率可将重复序列区分为高度重复序列(10 6次)、中度重复序列(10 3~10 4)及单拷贝序列。单拷贝序列在整个基因组中只出现一次或很少的几次。
(4)基因不连续性:真核结构基因两侧存在不被转录的非编码序列,往往是基因表达的调控区。在编码基因内部尚有一些不为蛋白质所编码的间隔序列,称内含子,而编码序列称为外显子,因此真核基因是不连续的。2、真核基因表达调控特点
(1)真核RNA聚合酶有三种,即RNA polⅠ、Ⅱ、Ⅲ,分别负责三种RNA转录,TATA盒结合蛋白为三种聚合酶所共有。
(2)活性染色体结构变化 :(1)对核酸酶敏感;(2)DNA拓扑结构变化。 天然状态的双链DNA以负性超螺旋的构象存在。当基因活化时,RNA聚合酶前方的转录区DNA拓扑结构为正性超螺旋,后面的DNA则为负超螺旋,正性超螺旋不仅阻碍核小体结构形成,而且促进组蛋白 H 2A·H 2B二聚体的释放,有利转录。(3)DNA碱基修饰变化。 在真核DNA有约5%的胞嘧啶被甲基化为5甲基胞嘧啶,甲基化范围与基因表达程度是反比关系。处于转录活化状态的基因CpG序列一般是低甲基化的。(4)组蛋白变化。 ①H 1样组蛋白减少。②H 2A·H 2B二聚体不稳定性增加。③组蛋白修饰:最常见的修饰有乙酰化、泛素化,修饰后使核小体结构变得不稳定。④H3组蛋白巯基暴露。
(3)正性调节占主导,提高了蛋白 -DNA相互作用的指导性,经济有效 ; (4)转录与翻译间隔进行,真核细胞有细胞核及胞浆等区间分布,转录与翻译在不同亚细胞结构中进行。(5)转录后修饰、加工 。3、原核与真核生物基因表达调控的比较
(1)共同点:
a 结构基因有调控序列;
b 表达的复杂性;
c 表达的时空性;
(2)不同点:
a 真核生物更复杂;
b 基因及基因组的结构特点;
c 转录与翻译的间断性;
d 转录后加工过程;
e 正负调控机制
f RNA聚合酶
五、基因与性状1、基本概念
基因:Gene
性状:Trait or Character
质量性状:Qualitative Trait
数量性状:Quantitative Trait
单基因控制性状:豌豆7对相对性状的遗传;
多基因控制性状:基因自由组合、连锁互换、基因互作。
一因多效:一个基因影响多个性状,即基因的多效性;
多因一效:多个基因控制一个性状 。
2、基因(型)控制性状
(1)作用途径:转录、翻译、生理生化反应(2) 基因(组)差异决定表型差异
DNA差异 表型差异
a. 重要功能基因的遗传效应;
b. 多基因间的综合作用
---数量性状的微效多基因假说(加性效应);
---基因间的复杂互作机制(非加性:显性、互补、上位、抑制、重叠等);
---基因表达调节蛋白,如转录调控因子。DNA(Gene) mRNA Protein Traitsc. 基因变异来源
---编码区变异直接影响产物的结构和功能;
---非编码区变异通过影响基因表达过程,调节合成产物量;
---大部分DNA变异表现为SNP。
d. 基因通过基因型发挥作用
---亲子相似性:1/2
---生命千姿百态:世界上找不到完全相同的两片树叶;
---性连锁基因,印记基因;(3)非DNA序列变异影响表型
a. 表(观)遗传学(Epigenetics)(表型~/外因~/发育~/拟~):研究基因型产生表型的过程;
b. 特点:可遗传;基因表达的改变;无DNA序列变化;
c. 遗传机制:
---DNA甲基化
---组蛋白修饰
---染色质改型
---RNA干涉。
《遗传》,2005.01
3、基因型与环境互作决定表型
基因(型)+环境=性状
(1)表现度:基因表现的程度,即在环境影响下基因在表现上的差异。
(2)环境的重要性:
a. 遗传、营养、生态环境的关系
(分子营养、分子生态)
b. 体细胞克隆:Hitler or Einstein
遗传信息的传递
一、DNA复制
二、DNA的转录
三、蛋白质的生物合成
四、基因表达调控
五、基因与性状 遗传信息的传递
主要内容遗传信息的传递概叙遗传信息的传递(中心法则)包括以下过程:DNA复制、DNA转录(逆转录、RNA复制)、蛋白质合成一
DNA复制
以亲代DNA分子为模板合成一个新的与亲代模板结构相同的子代DNA分子的过程。2.半不连续合成
DNA的双螺旋结构中的两条链是反向平行的,当复制开始解链时,亲代DNA分子中一条母链的方向为5′~3′,另一条母链的方向为3′~5′。DNA聚合酶只能催化5′~3′合成方向。在以3′~5′方向的母链为模板时,复制合成出一条5′~3′方向的前导链,前导链的前进方向与复制叉的行进方向一致,前导链的合成是连续进行的。而另一条母链仍以3′~5′方向作为模板,复制合成一条5′~3′方向的随从链,因此随从链合成方向是与复制叉的行进方向相反的。随从链的合成是不连续进行的,先合成许多片段,即冈崎片段。最后各冈崎片段再连接成为一条长链。由于前导链的合成连续进行的,而随从链的合成是不连续进行的,所以从总体上看DNA的复制是半不连续复制。(二)、 DNA复制所需的酶及蛋白质
1.DNA聚合酶
DNA复制时,以DNA为模板,以A、T、C、G为底物,按照碱基互补配对原则,沿5’→3’方向合成新的DNA双链。
原核生物DNA聚合酶包括:DNA聚合酶Ⅰ:具有三种功能—即5’→3’ 聚合酶活性、 3’→5’外切校正活性、 5’→3’外切酶活性; DNA聚合酶Ⅱ;DNA聚合酶Ⅲ: 细菌DNA复制的主要聚合酶,具有5’→3’ 聚合酶活性、 3’→5’外切校正活性。真核生物DNA聚合酶有α、β、γ、δ、ε 五种。2.引发酶
起始DNA复制。真核生物DNA聚合酶Ⅰ具有引发酶活性;原核生物由dnaG基因编码。
3 .DNA连接酶
将后随链中的各冈崎片段连接成一条完整的互补链。
4、拓扑异构酶
消除、维持、形成DNA的超螺旋结构,分Ⅰ、Ⅱ两种。 5、解链酶
解开DNA分子的互补双链作为复制的模板。
6、单链结合蛋白
结合解开的单链DNA,保护其不被水解和恢复DNA双链结构,又称为双螺旋反稳定蛋白。(三)、 DNA复制的过程
1.DNA复制起始:蛋白质-DNA复合物形成,复制叉及引发体产生。
2、DNA复制延伸:复制叉前移、前导链及后随链延伸。
3、DNA复制的终止:RNA引物切除、缺口补齐、冈崎片段连接。(四)、原核生物及真核生物DNA复制的比较
1.共同点:
(1)都具有半保留和半不连续特征;(2)都需要特定的酶和蛋白质;(3)复制过程都包括起始、延伸和终止三个阶段。
2、不同点:
(1)复制过程所需要的酶和蛋白因子不同;(2)原核生物DNA为环状分子,采用θ型复制和滚环式复制;真核生物通过端粒酶识别和结合端粒,维持线性DNA分子的恒定长度;(3)原核生物只有1个复制起点,复制速度快、冈崎片段长,能连续复制;真核生物有多个复制起点,复制速度慢、冈崎片段短,不能连续复制。二、DNA的转录 (Transcription)以DNA为模板,A、U、C、G为底物,按照碱基互补配对原则从5’→3’方向合成RNA的过程,称转录。2、真核生物的RNA聚合酶2.mRNA前体的加工
(1)5′末端帽子的生成
过程:一个m7G连接到mRNA的5‘端形成(脱Pi、甲基化),真核生物有0、1、2三种类型帽子结构。
功能:① 维持mRNA的一级结构,避免磷酸酶和核酶的攻击; ② 提供核糖体的识别部位。
(2)3′末端多聚A尾的生成
通过识别加尾信号AATAAA,在其下游20bp处加上100-200个A,形成polyA尾巴;
功能:①增加mRNA的稳定性;②方便mRNA从细胞核进入细胞质;③增强翻译效率;(3)剪接作用
在核酸内切酶作用下剪切掉内含子,然后在连接酶作用下,连接各部分外显子。
(4) RNA编辑
在转录产物中插入、删除或取代一些核苷酸残基,生成具有正确翻译功能的模板,此即所谓 RNA的编辑作用。 编辑过程由一个或多个小分子的“指导RNA”提供mRNA的编辑信息,并作为模板指导其进行编辑,在编辑体的帮助下进行编辑。(5)甲基化修饰
原核生物mRNA分子中不含有稀有碱基,但真核生物的mRNA中则含有甲基化核苷酸,除了在hnRNA的5′端帽子结构中含有2-3个甲基化核苷外;在分子内部还会有l-2个m6A存在于非编码区。在序列中, m6A总是位于胞苷之后,形成了…NC m6A N序列。 m6A的生成是在hnRNA的剪接作用之前发生的。
真核生物的CpG岛的C容易形成甲基化,常常突变成T。在基因的末端通常存在一些富含双核苷酸“CG”的区域,称为“CpG岛”(CpG island)。在人类基因组内,存在有近3万个CpG岛;在大多数染色体上,平均每100万碱基含有5~15个CpG岛,其中有1.8万多个CpG岛的GC含量为60%~70%。通常,这些CpG岛不仅是基因的一种标志,而且还参与基因表达的调控和影响染色质的结构。Epigenetics3.tRNA的转录后加工
①在核酸内切酶 RNaseP作用下,从5′末端切除多余的核苷酸;
②在核酸外切酶 RNaseD作用下,从3′末端切除多余的核苷酸;
③核苷酸转移酶催化, 3′末端加CCA-OH,为tRNA加I特有反应;
④核酸内切酶催化进行剪切反应,剪掉内含子,由连接酶连接外显子部分;
⑤ 化学修饰作用,如甲基化、脱氨基、还原反应。
4. rRNA的转录后加工
原核生物有 16S、23S及 5S三种 rRNA,这三种rRNA均存在于30S的rRNA前体中。转录作用完成后,在RNaseⅢ催化下,将rRNA前体切开产生16S、25S及 5S rRNA的中间前体。进一步在核酸酶的作用下,切去部分间隔序列,产生成熟的 16S、23S及5S rRNA,还有成熟的 tRNA。并对16S rRNA进行甲基化修饰,生成稀有碱基。与4S rRNA加I变化不大。
真核生物的核蛋白体中有18S、5.8S及5S rRNA。 5SrRNA自己独立成体系,在成熟过程中加工甚少,不进行修饰和剪切。 45S rRNA前体中包含有 18S、5.8S及 28SrRNA。在加工过程中,分子广泛地进行甲基化修饰,主要是在28S及18S中。甲基化作用多发生于核糖上,较少在碱基上。随后45 S rRNA前体经核酸酶顺序剪切下生成18S、5.8S、28S rRNA。
三、蛋白质的合成-翻译 (Translation) 基因的遗传信息在转录过程中从DNA转移到mRNA,再由mRNA将这种遗传信息表达为蛋白质中氨基酸顺序的过程叫做翻译,即蛋白质的合成。
(一)、 蛋白质合成体系
1、mRNA提供遗传密码
在mRNA中,每3个相互邻接的核苷酸,其特定排列顺序,在蛋白质生物合成中可被体现为某种氨基酸或合成的终止信号者称为密码子,统称遗传密码。它具有连续性、简并性、通用性。UAA、UAG、UGA为肽链的终止信号,不代表任何氨基酸,称为终止密码子;密码子AUG不仅代表一定氨基酸,而且位于mRNA启始部分,称起始密码子。编码同一氨基酸的称同义密码子。起编码氨基酸作用的称有义密码子。
遗传密码及相应的氨基酸2、tRNA转运氨基酸
tRNA上有由3个核苷酸组成的反密码子,与mRNA上的密码子按碱基互补配对原则结合, tRNA与对应氨基酸结合成为氨基酰tRNA,氨基酰tRNA才能准确的在mRNA上对号入座。
3、核糖(蛋白)体提供装配场所
核蛋白体相当于装配体,当核蛋白体在mRNA上每向前移动一个密码子的位置,肽链上即增加一个新的氨基酸,直至终止信号。
4、氨基酸为合成原料
20种氨基酸(二)、 蛋白质合成过程
1.启动阶段
在蛋白质生物合成的启动阶段,核蛋白体的大、小亚基,mRNA与一种具有启动作用的氨基酸tRNA共同构成启动复合体。
2.肽链延长阶段
根据 mRNA上密码子的要求,新的氨基酸不断相应的被特异的tRNA运至核蛋白体受位,形成肽键。
3.终止阶段
当多肽链合成已完成,并且“受位”上已出现终止信号,此后即转入终止阶段。终止阶段包括已合成完毕的肽链被水解释放,以及核蛋白体与tRNA从mRNA上脱落的过程。(三)、 翻译后加工
1.一级产物的修饰:(1)去除N-甲酰基或N-蛋氨酸;(2)个别氨基酸的修饰;(3)水解修饰;(4)切除部分肽链(连接肽、信号肽)。
2.高级结构的修饰:肽链释放后可自行根据其一级结构的特征折叠、盘曲成高级结构。此外,高级结构的修饰还包括(1)折叠、(2)亚基聚合、(3)辅基连接等行为。
3.蛋白质合成的靶向输送:蛋白质合成后,定向地到达其执行功能的目标地点,称为靶向输送;(2)切除。四、基因表达的调控(一)、 基因及其结构
1.基因概念的发展
(1)1866年孟德尔在《植物杂交试验》中提出的遗传因子概念,是基因雏形名词。 (2)1909年丹麦遗传学家约翰逊在《精密遗传学原理》中提出“基因”概念来替代孟德尔假定的“遗传因子”。
(3) 1926年摩尔根的巨著《基因论》出版,提出基因以直线形式排列,决定特定性状,能发生突变和交换,它不仅是决定性状的功能单位,而且是一个突变单位和交换单位。
(4) 1957年法国遗传学家本滋尔提出顺反子学说,认为基因是DNA分子上一段核苷酸顺序,负责着遗传信息传递。2.基因的几个相关概念
(1)基因:可转录一条完整的RNA分子或编码一条多肽链的一段DNA序列;产物具有一定的生物学功能;
(2)假基因(pseudo gene):同已知的基因相似,但位于不同位点,因缺失或突变而不能转录或翻译,是没有功能的基因。
(3)等位基因(allele):位于同源染色体上,位点相同,控制着同一性状的基因。
(4)结构基因(structural gene):可编码RNA或蛋白质的一段DNA序列。
(5)调控基因(regulator gene):其产物参与调控其他结构基因表达的基因。
(6)重叠基因{overlapping gene}:同一段DNA的编码顺序,由于阅读框架的不同或终止早晚的不同,同时编码两个或两个以上多肽链的基因。(7)隔裂基因(split gene):一个结构基因内部为一个或更多的不翻译的编码顺序,如内含子(intron)所隔裂的现象。
(8)跳跃基因(jumping gene):可作为插入因子和转座因子移动的DNA序列,也称转座因子。
(9)主基因(major gene):对于性状的作用比较明显,容易从杂种分离世代鉴别开来。
(10)修饰基因(modifying gene):一组效果微小的基因能增强或削弱主基因对表型的作用,这类微效基因在遗传学上称为修饰基因。
(11)管(持)家基因:某些基因是维持细胞基本代谢所必须的基因,其在一个生物个体的几乎所有细胞中持续表达,称为~;
(12)奢侈基因:有些基因在一些分化细胞中活动,是细胞分化、生物发育的基础,称为~。 3.基因的结构5‘侧翼区(5‘ –Flanking Region) :基因5‘端部到转录起始点间的部分;
5‘UTR: 5‘非翻译区(Un-translated Region);
启动子:指RNA聚合酶结合并起动转录的DNA序列,通常包括TATA框等结构;
转录起始点
转录终止点
起始密码子(Start Codon):通常为ATG;
终止密码子(Stop Codon):通常为TAA/G;
Poly(A)信号:决定加尾(转录终止)位置;
外显子(Exon):出现在成熟RNA中的有效DNA区段;
内含子(Intron):基因内部的部分序列并不出现在成熟mRNA中,这些间隔序列称为内含子, 符合“GT-AG”规则。真核生物基因的普通结构(二)、基因表达的概叙
1、概念
基因表达就是基因转录及翻译的过程。在一定调节机制控制下,大多数基因经历基因激活、转录及翻译等过程,产生具有特异生物学功能的蛋白质分子,但并非所有基因表达过程都产生蛋白质, tRNA、rRNA编码基因转录合成RNA的过程也属于基因表达。
2、基因表达的时间性及空间性
基因表达的时间、空间特异性由特异基因的启动子和增强子与调节蛋白相互作用决定。(1)时间特异性:按功能需要,某一特定基因的表达严格按特定的时间顺序发生,这是基因表达的时间特异性。 多细胞生物基因表达的时间特异性又称阶段特异性。(2)空间特异性:在个体生长全过程,某种基因产物在个体按不同组织空间顺序出现,这就是基因表达的空间特异性。又称细胞特异性或组织特异性。4、基因表达的复杂性表现为(1)多级调控:染色体和基因的结构活化、转录起始、转录后加工及转运、mRNA降解、翻译及翻译后加工及蛋白质降解等均为基因表达调控的控制点。(2)多要素综合作用:DNA序列、调节蛋白和RNA聚合酶等的结合作用。
5、基因表达的生物学意义:主要体现在适应环境,维持生长、增殖、个体发育和分化等方面。(三)、原核生物基因表达调控
1、操纵子学说
--原核生物的基因表达可在 DNA水平、转录水平和翻译水平三个层次进行调控,其中转录水平调控是基因表达调控的主要环节;
--操纵子是原核生物转录水平调控的主要方式,操纵子由调节基因R、操纵基因O和结构基因S组成;
--操纵子通过调节基因编码的调节蛋白开启或关闭操纵基因,调控结构基因的表达;
--如果调节蛋白关闭基因表达活性,称为负调控;反之,调节蛋白开启基因表达活性,称为正调控;
--一些小分子物质可作用于调节蛋白,开启转录活性的为诱导物,反之为辅阻遏物、超阻遏物。A转
乙
酰
酶?-
半
乳
糖
苷
透过酶ZY2、操纵子结构和功能
--乳糖操纵子模型;
--乳糖操纵子的结构;P’ R--乳糖操纵子的功能(负调控机制)3、操纵子的其他调控形式
(1)色氨酸操纵子;
(2)半乳糖操纵子(gal operon)
(3)阿拉伯糖操纵子(ara operon)
4、基因表达的时空调控
噬菌体SPO1侵染枯草杆菌:侵染后,早期基因立即转录;4-5m后,转为中期基因表达;8-12m后转为晚期基因表达。
5、DNA序列重排的调控
DNA序列的重排调控基因的表达,如鼠伤寒沙门氏菌的相转变机制(包括P的DNA倒位)。(四)、真核生物基因表达调控
1、真核基因组结构特点:基因组(Genome)是指一特定生物体的整套(单倍体)遗传物质的总和 。
(1)真核基因组结构庞大:哺乳类动物基因组DNA长达10 9个bp。
(2)单顺反子:真核基因转录产物为单顺反子,即一个编码基因转录生成一个 mRNA分子,经翻译生成一条多肽链。顺反子表示一个起作用的单位,其所包括的一段DNA与一个多肽链的合成相对应,是基因的基本功能和转录单位,一个基因可有几个顺反子,一个顺反子产生一条mRNA。
(3)重复序列:在原核、真核DNA中都有重复出现的核苷酸序列,但在真核更普遍。根据重复频率可将重复序列区分为高度重复序列(10 6次)、中度重复序列(10 3~10 4)及单拷贝序列。单拷贝序列在整个基因组中只出现一次或很少的几次。
(4)基因不连续性:真核结构基因两侧存在不被转录的非编码序列,往往是基因表达的调控区。在编码基因内部尚有一些不为蛋白质所编码的间隔序列,称内含子,而编码序列称为外显子,因此真核基因是不连续的。2、真核基因表达调控特点
(1)真核RNA聚合酶有三种,即RNA polⅠ、Ⅱ、Ⅲ,分别负责三种RNA转录,TATA盒结合蛋白为三种聚合酶所共有。
(2)活性染色体结构变化 :(1)对核酸酶敏感;(2)DNA拓扑结构变化。 天然状态的双链DNA以负性超螺旋的构象存在。当基因活化时,RNA聚合酶前方的转录区DNA拓扑结构为正性超螺旋,后面的DNA则为负超螺旋,正性超螺旋不仅阻碍核小体结构形成,而且促进组蛋白 H 2A·H 2B二聚体的释放,有利转录。(3)DNA碱基修饰变化。 在真核DNA有约5%的胞嘧啶被甲基化为5甲基胞嘧啶,甲基化范围与基因表达程度是反比关系。处于转录活化状态的基因CpG序列一般是低甲基化的。(4)组蛋白变化。 ①H 1样组蛋白减少。②H 2A·H 2B二聚体不稳定性增加。③组蛋白修饰:最常见的修饰有乙酰化、泛素化,修饰后使核小体结构变得不稳定。④H3组蛋白巯基暴露。
(3)正性调节占主导,提高了蛋白 -DNA相互作用的指导性,经济有效 ; (4)转录与翻译间隔进行,真核细胞有细胞核及胞浆等区间分布,转录与翻译在不同亚细胞结构中进行。(5)转录后修饰、加工 。3、原核与真核生物基因表达调控的比较
(1)共同点:
a 结构基因有调控序列;
b 表达的复杂性;
c 表达的时空性;
(2)不同点:
a 真核生物更复杂;
b 基因及基因组的结构特点;
c 转录与翻译的间断性;
d 转录后加工过程;
e 正负调控机制
f RNA聚合酶
五、基因与性状1、基本概念
基因:Gene
性状:Trait or Character
质量性状:Qualitative Trait
数量性状:Quantitative Trait
单基因控制性状:豌豆7对相对性状的遗传;
多基因控制性状:基因自由组合、连锁互换、基因互作。
一因多效:一个基因影响多个性状,即基因的多效性;
多因一效:多个基因控制一个性状 。
2、基因(型)控制性状
(1)作用途径:转录、翻译、生理生化反应(2) 基因(组)差异决定表型差异
DNA差异 表型差异
a. 重要功能基因的遗传效应;
b. 多基因间的综合作用
---数量性状的微效多基因假说(加性效应);
---基因间的复杂互作机制(非加性:显性、互补、上位、抑制、重叠等);
---基因表达调节蛋白,如转录调控因子。DNA(Gene) mRNA Protein Traitsc. 基因变异来源
---编码区变异直接影响产物的结构和功能;
---非编码区变异通过影响基因表达过程,调节合成产物量;
---大部分DNA变异表现为SNP。
d. 基因通过基因型发挥作用
---亲子相似性:1/2
---生命千姿百态:世界上找不到完全相同的两片树叶;
---性连锁基因,印记基因;(3)非DNA序列变异影响表型
a. 表(观)遗传学(Epigenetics)(表型~/外因~/发育~/拟~):研究基因型产生表型的过程;
b. 特点:可遗传;基因表达的改变;无DNA序列变化;
c. 遗传机制:
---DNA甲基化
---组蛋白修饰
---染色质改型
---RNA干涉。
《遗传》,2005.01
3、基因型与环境互作决定表型
基因(型)+环境=性状
(1)表现度:基因表现的程度,即在环境影响下基因在表现上的差异。
(2)环境的重要性:
a. 遗传、营养、生态环境的关系
(分子营养、分子生态)
b. 体细胞克隆:Hitler or Einstein