翻译
图片预览









文档简介
(共96张PPT)
第四章 从mRNA到蛋白质——翻译
第一节 mRNA与遗传密码
一、遗传密码
二、遗传密码的特性
一、遗传密码
mRNA是翻译的直接模板
遗传学将编码一个多肽的遗传单位称为顺反子(cistron)。
原核细胞中数个结构基因常串联为一个转录单位,转录生成的mRNA可编码几种功能相关的蛋白质,为多顺反子。
真核mRNA只编码一种蛋白质,为单顺反子。
多顺反子与单顺反子
mRNA上的遗传密码
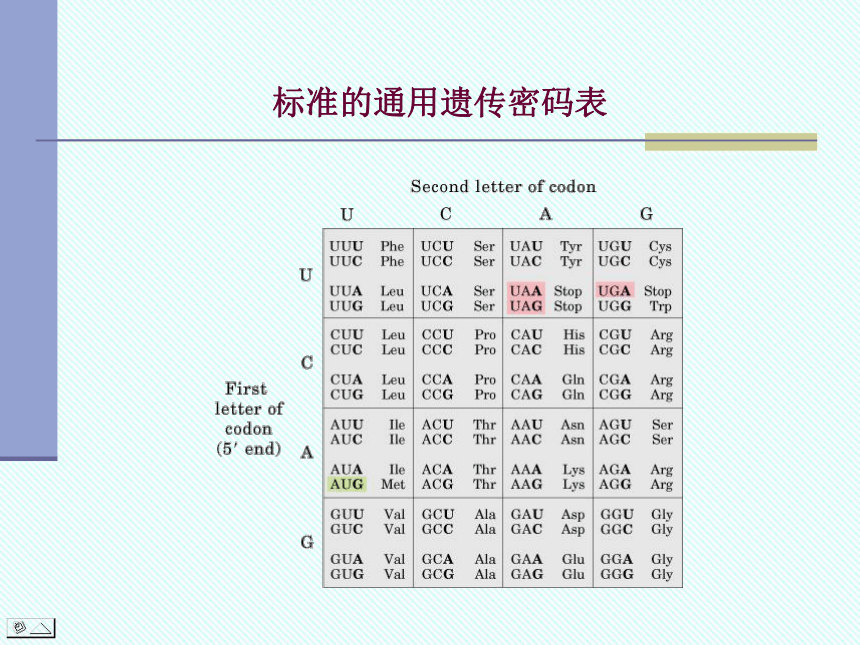
作为指导蛋白质生物合成的模板,mRNA中每三个相邻的核苷酸组成三联体,代表一个氨基酸的信息,此三联体就称为密码(codon)。共有64种不同的密码。其中:
起始密码(initiation codon): AUG
终止密码(termination codon):
UAA,UAG,UGA
标准的通用遗传密码表
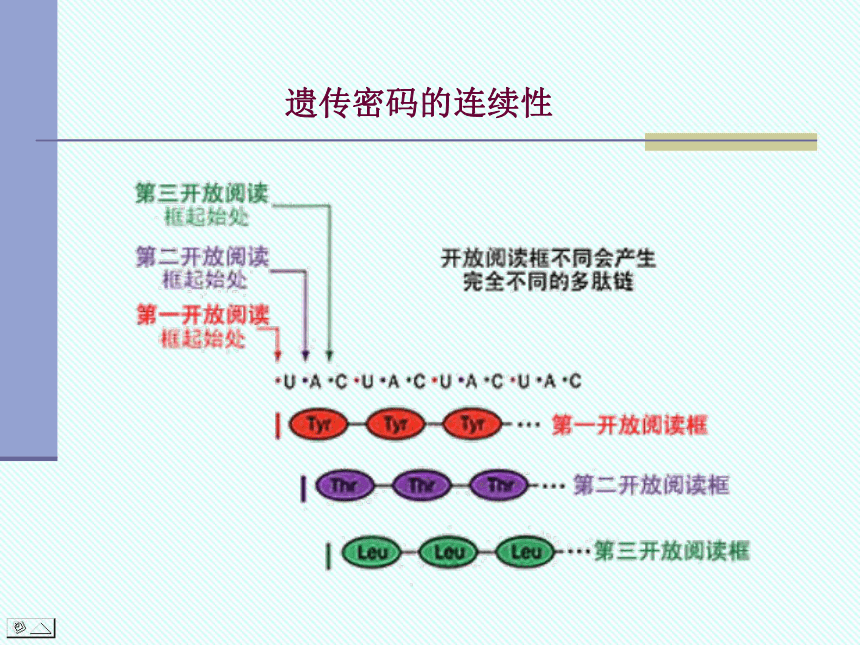
从mRNA 5 端起始密码子AUG到3 端终止密码子之间的核苷酸序列,各个三联体密码连续排列编码一个蛋白质多肽链,称为开放阅读框(open reading frame, ORF)。
1. 连续性(commaless)
指编码蛋白质氨基酸序列的各个三联体密码连续阅读。
二、遗传密码的特点
遗传密码的连续性
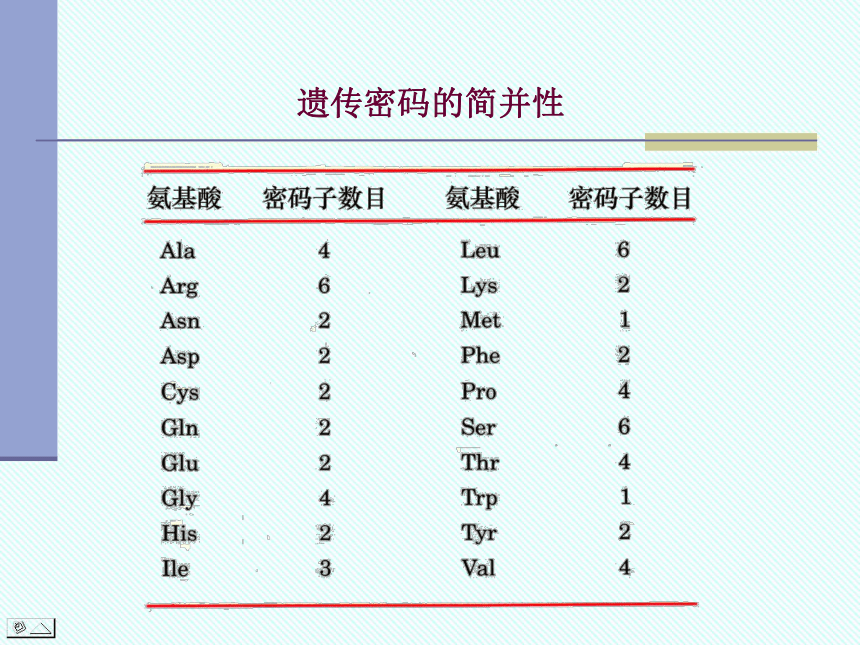
2. 简并性(degeneracy)
遗传密码中,除色氨酸和甲硫氨酸仅有一个密码子外,其余氨基酸有2、3、4个或多至6个三联体为其编码。
遗传密码的简并性在保持遗传稳定性上具有重要意义。
遗传密码的简并性
3. 通用性(universal)
蛋白质生物合成的整套密码,从原核生物到人类都通用。
已发现少数例外,如动物细胞的线粒体、植物细胞的叶绿体等。
密码的通用性进一步证明各种生物进化自同一祖先。
4. 方向性(direction)
指阅读mRNA模板上的三联体密码时,只能沿5’→3’方向进行。
5. 摆动性(wobble)
转运氨基酸的tRNA的反密码需要通过碱基互补与mRNA上的遗传密码反平行配对结合,但反密码与密码间不严格遵守常见的碱基配对规律,称为摆动配对。
密码子与反密码子的摆动配对
tRNA反密码子
第1位碱基 I U G A C
mRNA密码子
第3位碱基 U, C, A A, G U, C U G
U
摆动配对现象示意图
第二节 tRNA
DHU环
TψC环
附加环
反密码子环
aa接受臂
tRNA的三级结构示意图
第三节 rRNA与核糖体
一、rRNA与核糖体
二、核糖体的活性位点
核糖体是多肽链合成的场所,是由多种rRNA与蛋白质组装形成的复合体。
一、rRNA和核糖体
核蛋白体的组成
1.与模板mRNA和起始tRNA结合位点:主要与小亚基有关。
2.三个不同的tRNA结合位点:
⑴A位:又称受位或氨酰基位,可与新进入的氨基酰tRNA结合;由大、小亚基成分构成。
⑵P位:又称给位或肽酰基位,可与延伸中的肽酰基tRNA结合;由大、小亚基成分构成。
⑶E位:又称排出位,空载tRNA脱离核蛋白体前的结合位点;主要由大亚基成分构成。
二、核糖体的功能位点
3. 转肽酶活性:将给位上的肽酰基转移给受位上的氨基酰tRNA,形成肽键;由大亚基成分构成。
4. GTPase活性:水解GTP,获得能量;分别由大、小亚基成分构成。
5. 起动因子、延长因子及释放因子的结合位点:分别由大、小亚基成分构成。
原核生物翻译过程中核蛋白体结构模式
A位:氨基酰位
(aminoacyl site)
P位:肽酰位
(peptidyl site)
E位:排出位
(exit site)
多聚核糖体示意图
电镜下的多聚核糖体
第四节 蛋白质的合成机制
一、氨基酸的活化与携带
二、翻译起始
三、翻译延伸
四、翻译终止
蛋白质生物合成过程包括三个环节:
-氨基酸的活化与携带;
-多肽链的形成与延伸(翻译起始,翻译延伸,翻译终止);
-多肽链合成后的加工修饰。
一、氨基酸的活化与搬运
氨基酸的活化与携带反应由氨酰tRNA合成酶催化。
特定的tRNA与相应的氨基酸结合,生成氨酰tRNA,从而由tRNA携带活化的氨基酸参与蛋白质的生物合成。
氨基酸 + tRNA
氨基酰- tRNA
ATP
AMP+PPi
氨基酰-tRNA合成酶
氨基酰tRNA合成酶催化的反应:
氨基酸 +ATP-E → 氨基酰-AMP-E + PPi
第一步:活化反应
第二步:连接反应
氨基酰-AMP-E
+
tRNA
↓
氨基酰-tRNA
+
AMP
+
E
氨基酰-tRNA合成酶对底物氨基酸和tRNA都有高度特异性。
氨基酰-tRNA合成酶具有校正活性(proof-reading activity) 。
氨基酰-tRNA的表示方法:
Ala-tRNAAla
Ser-tRNASer
Met-tRNAMet
包括以下几个步骤:
核糖体大小亚基分离;
mRNA与小亚基定位结合;
起始氨基酰-tRNA的结合;
核糖体大亚基结合,翻译起始复合物形成。
二、翻译起始
(一)原核生物翻译起始复合物形成
IF-3
IF-1
1. 核糖体大、小亚基分离:
翻译起始因子IF-1和IF-3与小亚基结合,促进核糖体大、小亚基拆离,为新一轮合成作准备。
A
U
G
5'
3'
IF-3
IF-1
2. mRNA在小亚基的精确定位结合:
S-D序列
原核mRNA的起始部位由一段富含嘌呤的特殊核苷酸顺序组成,称为S-D序列(核糖体结合位点,RBS)。
16S rRNA存在一段富含嘧啶的序列,二者之间可通过碱基配对结合。
在mRNA上紧接S-D序列之后的序列,可被核糖体小亚基蛋白辨认结合。
IF-2
GTP
3. 起始氨基酰tRNA( fMet-tRNAimet )结合到小亚基
IF-3
IF-1
A
U
G
5'
3'
起始 fMet-tRNAimet以及IF2-GTP一起,识别结合小亚基P位,并对应模板mRNA的起始密码AUG。
IF-3
IF-1
IF-2
-GTP
-GDP
Pi
4. 核糖体大亚基结合,起始复合体形成:
A
U
G
5'
3'
50S大亚基与30S小亚基、模板mRNA结合, IF2结合的GTP被水解,三种IF脱离。核糖体,mRNA与起始fMet-tRNAifMet构成起始复合体。
(二)真核生物翻译起始复合体形成
真核生物翻译起始复合体的形成过程与原核生物类似,但参与的蛋白因子更多。
真核生物mRNA没有SD序列,帽子结构是翻译起始因子识别的对象
延伸方向从N端向C端
多肽链合成的延伸循环反应过程来完成
核糖体循环包括多肽链合成的氨酰tRNA结合(进位),肽键的形成(转肽)和移位(转位)三步反应。
三、肽链的延伸
1. 氨酰tRNA结合
与mRNA下一个密码相对应的氨基酰tRNA进入核糖体的A位。此步骤需GTP,Mg2+,和延伸因子EF-Tu参与。
EF-Ts帮助EF-Tu与GTP结合
2. 肽键的形成(peptide bond formation):
是由肽基转移酶催化的肽键形成过程,将P位上的tRNA所携带的甲酰蛋氨酰基或肽酰基转移到A位上的氨基酰tRNA上,与其氨基缩合形成肽键。
成肽反应过程
3. 移位(translocation):
核糖体向mRNA的3’侧移动相当于一个密码的距离,A位对应于新密码子。
肽酰基tRNA从A位移到P位。
去氨酰-tRNA 被挤入E位
需要延长因子EF-G,有转位酶活性,可结合并水解1分子GTP。
移位反应过程
进位
移位
转肽
核糖体循环的反应过程
核糖体沿mRNA链滑动,不断使多肽链延长,直到终止密码子进入A位。
1.识别:释放因子RF识别终止密码,进入核糖体的A位。
2.水解:RF使肽基转移酶变为酯酶,多肽链与tRNA之间的酯键被水解,多肽链释放。
3. 脱离:模板mRNA、释放因子RF以及空载tRNA与核糖体脱离。
RF因子有GTP水解酶活性,水解GTP提供能量
四、翻译终止
多肽链合成的终止过程
第五节 蛋白质的折叠、加工与运输
一、蛋白质的折叠
二、蛋白质的加工
三、蛋白质合成后的靶向输送
从核糖体释放出的新生多肽链不具备蛋白质生物活性,必需经过不同的翻译后复杂加工过程才转变为天然构象的功能蛋白。
主要包括:
多肽链折叠为天然的三维结构肽链;
一级结构的修饰;
高级结构修饰。
多肽链自身氨基酸顺序储存着蛋白质折叠的信息,即一级结构是空间构象的基础。
细胞中大多数天然蛋白质折叠需要其他酶、蛋白辅助。
新生肽链的折叠在肽链合成中、合成后完成,新生肽链N端在核糖体上一出现,肽链的折叠即开始。可随着序列的不断延伸肽链逐步折叠,产生正确的二级结构、结构域到形成完整的空间构象。
一、蛋白质的折叠
几种有促进蛋白折叠功能的大分子
分子伴侣 (molecular chaperon)
蛋白二硫键异构酶
肽-脯氨酰顺反异构酶
1. 分子伴侣:
分子伴侣是细胞一类保守蛋白质,可识别肽链的非天然构象,促进各功能域和整体蛋白质的正确折叠。
热休克蛋白(heat shock protein, HSP)
HSP90、HSP70、HSP40等。
HSP蛋白家族大多有ATP酶活性,利用ATP水解提供能力打开错误形成的次级键,促进重新折叠
2. 蛋白二硫键异构酶:
多肽链内或肽链之间二硫键的正确形成对稳定分泌蛋白、膜蛋白等的天然构象十分重要。
二硫键异构酶可在较大区段肽链中催化错配二硫键断裂并形成正确二硫键连接,最终使蛋白质形成热力学最稳定的天然构象。
3. 肽-脯氨酰顺反异构酶:
多肽链中肽酰-脯氨酸间形成的肽键有顺反两种异构体,空间构象明显差别。
肽酰-脯氨酰顺反异构酶可促进上述顺反两种异构体之间的转换。
N端甲酰蛋氨酸或蛋氨酸残基,往往在多肽合成完成之前被切除。
二、蛋白质的加工
① 去甲酰化:
甲酰化酶
甲酰蛋氨酸-肽
甲酸 + 蛋氨酸-肽
② 去蛋氨酰基:
蛋氨酸氨基肽酶
蛋氨酰-肽
蛋氨酸 + 肽
(一)N端甲酰蛋氨酸或蛋氨酸的切除
由专一性的蛋白酶催化,将部分肽段切除。
(二)氨基酸的共价修饰
由专一性的酶催化进行修饰,包括糖基化、磷酸化、甲酰化、甲基化、羟基化等。
(三)切除新生肽链中的非功能片段
胰岛素的加工过程
蛋白质合成后,定向地被输送到其执行功能的场所称为靶向输送(protein targeting)。
大多数情况下,被输送的蛋白质分子需穿过膜性结构,才能到达特定的地点。
所有靶向输送的蛋白质结构中存在分选信号,可引导蛋白质转移到细胞的适当靶部位,这一序列称为信号序列(signal sequence)。
三、蛋白质合成后的靶向输送
(一)分泌型蛋白的靶向输送
真核细胞分泌型蛋白前体合成后的靶向输送过程首先要进入内质网。
分泌型蛋白新生多肽链的N端的15~36个氨基酸残基的肽段, 有保守的氨基酸序列,引导蛋白质进入内质网,称为信号肽(signal peptide) 。
1. 信号肽与信号识别蛋白:
常见的信号肽由10~40个氨基酸残基组成,可分为三段。N端为带正电荷的氨基酸残基,中间为疏水的核心区,而C端由小分子氨基酸残基组成。在蛋白质被分泌后,信号肽序列可被信号肽酶识别并裂解。
信号识别蛋白(SRP):6种多肽和1个7S RNA组成,与信号肽结合,可导致蛋白质合成暂停。
常见分泌型蛋白质的信号肽序列
2. 分泌型蛋白进入内质网的过程:
分泌型蛋白的定向输送,是靠信号肽与胞浆中的信号识别蛋白(SRP)识别并特异结合,然后再通过SRP与内质网膜上的SRP受体(DP 停靠蛋白)识别并结合后,从而使核糖体结合到内质网膜上的核糖体受体上,新生肽链通过膜上通道穿越内质网膜,合成后的蛋白质进入内质网腔,最终通过膜泡运输将所携带的蛋白质送出细胞。
信号肽引导分泌型蛋白进入内质网
(二)线粒体蛋白的靶向输送
(三)核定位蛋白的靶向输送
核定位蛋白如何需要通过核孔,能引导蛋白质入核的信号序列我们称为核定位序列(NLS),核定位序列可位于核蛋白任何位置,不被切除。
入核过程:核转运因子(importin)识别核定位序列,并与之结合,形成复合物,停靠到核孔处,依靠Ran蛋白GTP酶水解活性水解GTP提供能量进入细胞核。入核后,复合物解离,核转运因子出核。
遗传密码的特点
tRNA的二级结构与高级结构
氨基酸的活化与氨酰tRNA合成酶
核糖体的结构与组成
原核与真核细胞翻译的起始过程
翻译延伸过程的循环反应
翻译终止过程
多肽链的折叠与参与的其他蛋白质
蛋白质的靶向运输机制
总结:真核原核翻译机制的差异
本章要点
1,DNA双链中,指导合成RNA的那条链称作
A 编码链
B 有义链
C 模板链
D 以上都不对
2,原核生物中,识别并结合启动子的是
A 因子
B 亚基
C 亚基
D 因子
3,参与原核生物转录延长的酶是
A 引物酶
B RNA聚合酶核心酶
C RNA聚合酶全酶
D RNA聚合酶II
4,真核生物中,mRNA由下面哪种酶转录
A RNA聚合酶I
B RNA聚合酶II
C RNA聚合酶III
D RNA酶
5,下面关于真核生物mRNA转录起始的说法错误的是
A 需要多种转录因子参与
B TFIID识别TATA盒
C 需要CTD被磷酸化才进入延伸阶段
D 转录从TATA盒开始
6,下面关于真核生物RNA聚合酶III的说法错误的是
A 合成tRNA
B 合成5S rRNA
C 启动子位于编码基因转录区内
D 合成5.8S rRNA
7,下面参与RNA剪接的RNA是:
A HnRNA
B snRNA
C 5SRNA
D tRNA
8,rRNA剪接过程中发挥作用的主要是:
A 核酶
B snRNA
C 核酸内切酶
D 分子伴侣
9,gRNA在以下哪个过程中发挥作用:
A DNA复制
B 转录
C RNA剪接
D RNA编辑
10,对于依赖蛋白质因子的终止机制的叙述错误的是:
A 终止子结构与不依赖与蛋白质因子的终止子有很大差异
B 需要 因子的参与
C 蛋白质因子最初结合于mRNA5’端
D 可能出现抗终止现象
11,下面事件在真核转录不发生而在原核转录中发生的是
A 戴帽
B 加尾
C 内含子的去除
D 转录过程中核糖体结合mRNA进行翻译
12,DNA分子中,转录的模板链被称为
A 随从链
B 先导链
C 编码链
D 无义链
13,关于真核生物mRNA的聚腺苷酸尾巴,下面说法正确的是
A 由模板DNA上聚T序列转录而成
B 转录结束后在初级转录产物的3’端直接加上
C 是翻译起始因子识别的位点
D 在细胞核内添加
14,哪种RNA具有较多的稀有核苷酸
A tRNA
B rRNA
C mRNA
D hnRNA
15,有氨基酸对应的密码子有多少个?
A 64
B 61
C 60
D 20
16,与tRNA上反密码子5’UAG 3’配对的密码子是
A 5'AUC 3'
B 5'UAG 3'
C 5'CUA 3'
D 5'GAU 3'
17,下列哪组密码子是终止密码子
A AAC,ACA,ACU
B AUG,AGU,GAU
C CAA,GAA,UAA
D UAA,UAG,UGA
18,密码子的摆动性表现在
A 反密码子第一位与密码子第三位配对不严格
B 反密码子第三位与密码子第一位配对不严格
C 反密码子第二位与密码子第二位配对不严格
D 反密码子第三位与密码子第三位配对不严格
19,tRNA高级结构为倒L型,其中L长臂的末端是
A 反密码子环
B 氨基酸臂
C 二氢尿嘧啶环
D 附加环
20,真核生物中,参与形成小亚基的rRNA是
A 28S rRNA
B 18S rRNA
C 16S rRNA
D 5S rRNA
21,有关蛋白质合成,下列哪条描述是错误的:
A 基本原料是20种氨基酸
B 直接模板是mRNA
C 合成的方向是从羧基端到氨基端
D 是一个多因子参加的耗能过程
22,下面关于氨酰tRNA合成酶的叙述错误的是
A 催化tRNA3’的羟基与氨基酸羧基形成脂键
B 对氨基酸有高度特异性,对tRNA没有特异性
C 有校正活性
D 活化过程中ATP被水解为AMP
23,下面哪个过程不消耗GTP
A 翻译起始因子释放
B 转肽
C 移位
D 肽链释放
24,翻译起始复合物的组成
A 核糖体+翻译起始因子
B 核糖体+翻译起始因子+mRNA
C 核糖体+mRNA+氨基酰tRNA
D 核糖体+mRNA+甲硫氨酰tRNA
25,翻译起始时与甲酰甲硫氨酰tRNA结合的起始因子是:
A IF1
B IF2
C IF3
D EF-Tu
26,关于移位,错误的是
A 需要EF-G蛋白因子
B 转位完成后,A位空留
C 肽酰tRNA从P位进入A位
D 核糖体延mRNA移动
27,下面关于转肽反应的叙述正确的是
A 肽链从A位tRNA上转移到P位tRNA上
B 需要肽基转移酶的催化
C 转肽反应发生位置在核糖体小亚单位上
D 转肽反应完成后,A位上的tRNA将离开核糖体
28,原核细胞与真核细胞肽链终止因子的数量分别是
A 3,多个
B 1,3
C 3,1
D 1,多个
第四章 从mRNA到蛋白质——翻译
第一节 mRNA与遗传密码
一、遗传密码
二、遗传密码的特性
一、遗传密码
mRNA是翻译的直接模板
遗传学将编码一个多肽的遗传单位称为顺反子(cistron)。
原核细胞中数个结构基因常串联为一个转录单位,转录生成的mRNA可编码几种功能相关的蛋白质,为多顺反子。
真核mRNA只编码一种蛋白质,为单顺反子。
多顺反子与单顺反子
mRNA上的遗传密码
作为指导蛋白质生物合成的模板,mRNA中每三个相邻的核苷酸组成三联体,代表一个氨基酸的信息,此三联体就称为密码(codon)。共有64种不同的密码。其中:
起始密码(initiation codon): AUG
终止密码(termination codon):
UAA,UAG,UGA
标准的通用遗传密码表
从mRNA 5 端起始密码子AUG到3 端终止密码子之间的核苷酸序列,各个三联体密码连续排列编码一个蛋白质多肽链,称为开放阅读框(open reading frame, ORF)。
1. 连续性(commaless)
指编码蛋白质氨基酸序列的各个三联体密码连续阅读。
二、遗传密码的特点
遗传密码的连续性
2. 简并性(degeneracy)
遗传密码中,除色氨酸和甲硫氨酸仅有一个密码子外,其余氨基酸有2、3、4个或多至6个三联体为其编码。
遗传密码的简并性在保持遗传稳定性上具有重要意义。
遗传密码的简并性
3. 通用性(universal)
蛋白质生物合成的整套密码,从原核生物到人类都通用。
已发现少数例外,如动物细胞的线粒体、植物细胞的叶绿体等。
密码的通用性进一步证明各种生物进化自同一祖先。
4. 方向性(direction)
指阅读mRNA模板上的三联体密码时,只能沿5’→3’方向进行。
5. 摆动性(wobble)
转运氨基酸的tRNA的反密码需要通过碱基互补与mRNA上的遗传密码反平行配对结合,但反密码与密码间不严格遵守常见的碱基配对规律,称为摆动配对。
密码子与反密码子的摆动配对
tRNA反密码子
第1位碱基 I U G A C
mRNA密码子
第3位碱基 U, C, A A, G U, C U G
U
摆动配对现象示意图
第二节 tRNA
DHU环
TψC环
附加环
反密码子环
aa接受臂
tRNA的三级结构示意图
第三节 rRNA与核糖体
一、rRNA与核糖体
二、核糖体的活性位点
核糖体是多肽链合成的场所,是由多种rRNA与蛋白质组装形成的复合体。
一、rRNA和核糖体
核蛋白体的组成
1.与模板mRNA和起始tRNA结合位点:主要与小亚基有关。
2.三个不同的tRNA结合位点:
⑴A位:又称受位或氨酰基位,可与新进入的氨基酰tRNA结合;由大、小亚基成分构成。
⑵P位:又称给位或肽酰基位,可与延伸中的肽酰基tRNA结合;由大、小亚基成分构成。
⑶E位:又称排出位,空载tRNA脱离核蛋白体前的结合位点;主要由大亚基成分构成。
二、核糖体的功能位点
3. 转肽酶活性:将给位上的肽酰基转移给受位上的氨基酰tRNA,形成肽键;由大亚基成分构成。
4. GTPase活性:水解GTP,获得能量;分别由大、小亚基成分构成。
5. 起动因子、延长因子及释放因子的结合位点:分别由大、小亚基成分构成。
原核生物翻译过程中核蛋白体结构模式
A位:氨基酰位
(aminoacyl site)
P位:肽酰位
(peptidyl site)
E位:排出位
(exit site)
多聚核糖体示意图
电镜下的多聚核糖体
第四节 蛋白质的合成机制
一、氨基酸的活化与携带
二、翻译起始
三、翻译延伸
四、翻译终止
蛋白质生物合成过程包括三个环节:
-氨基酸的活化与携带;
-多肽链的形成与延伸(翻译起始,翻译延伸,翻译终止);
-多肽链合成后的加工修饰。
一、氨基酸的活化与搬运
氨基酸的活化与携带反应由氨酰tRNA合成酶催化。
特定的tRNA与相应的氨基酸结合,生成氨酰tRNA,从而由tRNA携带活化的氨基酸参与蛋白质的生物合成。
氨基酸 + tRNA
氨基酰- tRNA
ATP
AMP+PPi
氨基酰-tRNA合成酶
氨基酰tRNA合成酶催化的反应:
氨基酸 +ATP-E → 氨基酰-AMP-E + PPi
第一步:活化反应
第二步:连接反应
氨基酰-AMP-E
+
tRNA
↓
氨基酰-tRNA
+
AMP
+
E
氨基酰-tRNA合成酶对底物氨基酸和tRNA都有高度特异性。
氨基酰-tRNA合成酶具有校正活性(proof-reading activity) 。
氨基酰-tRNA的表示方法:
Ala-tRNAAla
Ser-tRNASer
Met-tRNAMet
包括以下几个步骤:
核糖体大小亚基分离;
mRNA与小亚基定位结合;
起始氨基酰-tRNA的结合;
核糖体大亚基结合,翻译起始复合物形成。
二、翻译起始
(一)原核生物翻译起始复合物形成
IF-3
IF-1
1. 核糖体大、小亚基分离:
翻译起始因子IF-1和IF-3与小亚基结合,促进核糖体大、小亚基拆离,为新一轮合成作准备。
A
U
G
5'
3'
IF-3
IF-1
2. mRNA在小亚基的精确定位结合:
S-D序列
原核mRNA的起始部位由一段富含嘌呤的特殊核苷酸顺序组成,称为S-D序列(核糖体结合位点,RBS)。
16S rRNA存在一段富含嘧啶的序列,二者之间可通过碱基配对结合。
在mRNA上紧接S-D序列之后的序列,可被核糖体小亚基蛋白辨认结合。
IF-2
GTP
3. 起始氨基酰tRNA( fMet-tRNAimet )结合到小亚基
IF-3
IF-1
A
U
G
5'
3'
起始 fMet-tRNAimet以及IF2-GTP一起,识别结合小亚基P位,并对应模板mRNA的起始密码AUG。
IF-3
IF-1
IF-2
-GTP
-GDP
Pi
4. 核糖体大亚基结合,起始复合体形成:
A
U
G
5'
3'
50S大亚基与30S小亚基、模板mRNA结合, IF2结合的GTP被水解,三种IF脱离。核糖体,mRNA与起始fMet-tRNAifMet构成起始复合体。
(二)真核生物翻译起始复合体形成
真核生物翻译起始复合体的形成过程与原核生物类似,但参与的蛋白因子更多。
真核生物mRNA没有SD序列,帽子结构是翻译起始因子识别的对象
延伸方向从N端向C端
多肽链合成的延伸循环反应过程来完成
核糖体循环包括多肽链合成的氨酰tRNA结合(进位),肽键的形成(转肽)和移位(转位)三步反应。
三、肽链的延伸
1. 氨酰tRNA结合
与mRNA下一个密码相对应的氨基酰tRNA进入核糖体的A位。此步骤需GTP,Mg2+,和延伸因子EF-Tu参与。
EF-Ts帮助EF-Tu与GTP结合
2. 肽键的形成(peptide bond formation):
是由肽基转移酶催化的肽键形成过程,将P位上的tRNA所携带的甲酰蛋氨酰基或肽酰基转移到A位上的氨基酰tRNA上,与其氨基缩合形成肽键。
成肽反应过程
3. 移位(translocation):
核糖体向mRNA的3’侧移动相当于一个密码的距离,A位对应于新密码子。
肽酰基tRNA从A位移到P位。
去氨酰-tRNA 被挤入E位
需要延长因子EF-G,有转位酶活性,可结合并水解1分子GTP。
移位反应过程
进位
移位
转肽
核糖体循环的反应过程
核糖体沿mRNA链滑动,不断使多肽链延长,直到终止密码子进入A位。
1.识别:释放因子RF识别终止密码,进入核糖体的A位。
2.水解:RF使肽基转移酶变为酯酶,多肽链与tRNA之间的酯键被水解,多肽链释放。
3. 脱离:模板mRNA、释放因子RF以及空载tRNA与核糖体脱离。
RF因子有GTP水解酶活性,水解GTP提供能量
四、翻译终止
多肽链合成的终止过程
第五节 蛋白质的折叠、加工与运输
一、蛋白质的折叠
二、蛋白质的加工
三、蛋白质合成后的靶向输送
从核糖体释放出的新生多肽链不具备蛋白质生物活性,必需经过不同的翻译后复杂加工过程才转变为天然构象的功能蛋白。
主要包括:
多肽链折叠为天然的三维结构肽链;
一级结构的修饰;
高级结构修饰。
多肽链自身氨基酸顺序储存着蛋白质折叠的信息,即一级结构是空间构象的基础。
细胞中大多数天然蛋白质折叠需要其他酶、蛋白辅助。
新生肽链的折叠在肽链合成中、合成后完成,新生肽链N端在核糖体上一出现,肽链的折叠即开始。可随着序列的不断延伸肽链逐步折叠,产生正确的二级结构、结构域到形成完整的空间构象。
一、蛋白质的折叠
几种有促进蛋白折叠功能的大分子
分子伴侣 (molecular chaperon)
蛋白二硫键异构酶
肽-脯氨酰顺反异构酶
1. 分子伴侣:
分子伴侣是细胞一类保守蛋白质,可识别肽链的非天然构象,促进各功能域和整体蛋白质的正确折叠。
热休克蛋白(heat shock protein, HSP)
HSP90、HSP70、HSP40等。
HSP蛋白家族大多有ATP酶活性,利用ATP水解提供能力打开错误形成的次级键,促进重新折叠
2. 蛋白二硫键异构酶:
多肽链内或肽链之间二硫键的正确形成对稳定分泌蛋白、膜蛋白等的天然构象十分重要。
二硫键异构酶可在较大区段肽链中催化错配二硫键断裂并形成正确二硫键连接,最终使蛋白质形成热力学最稳定的天然构象。
3. 肽-脯氨酰顺反异构酶:
多肽链中肽酰-脯氨酸间形成的肽键有顺反两种异构体,空间构象明显差别。
肽酰-脯氨酰顺反异构酶可促进上述顺反两种异构体之间的转换。
N端甲酰蛋氨酸或蛋氨酸残基,往往在多肽合成完成之前被切除。
二、蛋白质的加工
① 去甲酰化:
甲酰化酶
甲酰蛋氨酸-肽
甲酸 + 蛋氨酸-肽
② 去蛋氨酰基:
蛋氨酸氨基肽酶
蛋氨酰-肽
蛋氨酸 + 肽
(一)N端甲酰蛋氨酸或蛋氨酸的切除
由专一性的蛋白酶催化,将部分肽段切除。
(二)氨基酸的共价修饰
由专一性的酶催化进行修饰,包括糖基化、磷酸化、甲酰化、甲基化、羟基化等。
(三)切除新生肽链中的非功能片段
胰岛素的加工过程
蛋白质合成后,定向地被输送到其执行功能的场所称为靶向输送(protein targeting)。
大多数情况下,被输送的蛋白质分子需穿过膜性结构,才能到达特定的地点。
所有靶向输送的蛋白质结构中存在分选信号,可引导蛋白质转移到细胞的适当靶部位,这一序列称为信号序列(signal sequence)。
三、蛋白质合成后的靶向输送
(一)分泌型蛋白的靶向输送
真核细胞分泌型蛋白前体合成后的靶向输送过程首先要进入内质网。
分泌型蛋白新生多肽链的N端的15~36个氨基酸残基的肽段, 有保守的氨基酸序列,引导蛋白质进入内质网,称为信号肽(signal peptide) 。
1. 信号肽与信号识别蛋白:
常见的信号肽由10~40个氨基酸残基组成,可分为三段。N端为带正电荷的氨基酸残基,中间为疏水的核心区,而C端由小分子氨基酸残基组成。在蛋白质被分泌后,信号肽序列可被信号肽酶识别并裂解。
信号识别蛋白(SRP):6种多肽和1个7S RNA组成,与信号肽结合,可导致蛋白质合成暂停。
常见分泌型蛋白质的信号肽序列
2. 分泌型蛋白进入内质网的过程:
分泌型蛋白的定向输送,是靠信号肽与胞浆中的信号识别蛋白(SRP)识别并特异结合,然后再通过SRP与内质网膜上的SRP受体(DP 停靠蛋白)识别并结合后,从而使核糖体结合到内质网膜上的核糖体受体上,新生肽链通过膜上通道穿越内质网膜,合成后的蛋白质进入内质网腔,最终通过膜泡运输将所携带的蛋白质送出细胞。
信号肽引导分泌型蛋白进入内质网
(二)线粒体蛋白的靶向输送
(三)核定位蛋白的靶向输送
核定位蛋白如何需要通过核孔,能引导蛋白质入核的信号序列我们称为核定位序列(NLS),核定位序列可位于核蛋白任何位置,不被切除。
入核过程:核转运因子(importin)识别核定位序列,并与之结合,形成复合物,停靠到核孔处,依靠Ran蛋白GTP酶水解活性水解GTP提供能量进入细胞核。入核后,复合物解离,核转运因子出核。
遗传密码的特点
tRNA的二级结构与高级结构
氨基酸的活化与氨酰tRNA合成酶
核糖体的结构与组成
原核与真核细胞翻译的起始过程
翻译延伸过程的循环反应
翻译终止过程
多肽链的折叠与参与的其他蛋白质
蛋白质的靶向运输机制
总结:真核原核翻译机制的差异
本章要点
1,DNA双链中,指导合成RNA的那条链称作
A 编码链
B 有义链
C 模板链
D 以上都不对
2,原核生物中,识别并结合启动子的是
A 因子
B 亚基
C 亚基
D 因子
3,参与原核生物转录延长的酶是
A 引物酶
B RNA聚合酶核心酶
C RNA聚合酶全酶
D RNA聚合酶II
4,真核生物中,mRNA由下面哪种酶转录
A RNA聚合酶I
B RNA聚合酶II
C RNA聚合酶III
D RNA酶
5,下面关于真核生物mRNA转录起始的说法错误的是
A 需要多种转录因子参与
B TFIID识别TATA盒
C 需要CTD被磷酸化才进入延伸阶段
D 转录从TATA盒开始
6,下面关于真核生物RNA聚合酶III的说法错误的是
A 合成tRNA
B 合成5S rRNA
C 启动子位于编码基因转录区内
D 合成5.8S rRNA
7,下面参与RNA剪接的RNA是:
A HnRNA
B snRNA
C 5SRNA
D tRNA
8,rRNA剪接过程中发挥作用的主要是:
A 核酶
B snRNA
C 核酸内切酶
D 分子伴侣
9,gRNA在以下哪个过程中发挥作用:
A DNA复制
B 转录
C RNA剪接
D RNA编辑
10,对于依赖蛋白质因子的终止机制的叙述错误的是:
A 终止子结构与不依赖与蛋白质因子的终止子有很大差异
B 需要 因子的参与
C 蛋白质因子最初结合于mRNA5’端
D 可能出现抗终止现象
11,下面事件在真核转录不发生而在原核转录中发生的是
A 戴帽
B 加尾
C 内含子的去除
D 转录过程中核糖体结合mRNA进行翻译
12,DNA分子中,转录的模板链被称为
A 随从链
B 先导链
C 编码链
D 无义链
13,关于真核生物mRNA的聚腺苷酸尾巴,下面说法正确的是
A 由模板DNA上聚T序列转录而成
B 转录结束后在初级转录产物的3’端直接加上
C 是翻译起始因子识别的位点
D 在细胞核内添加
14,哪种RNA具有较多的稀有核苷酸
A tRNA
B rRNA
C mRNA
D hnRNA
15,有氨基酸对应的密码子有多少个?
A 64
B 61
C 60
D 20
16,与tRNA上反密码子5’UAG 3’配对的密码子是
A 5'AUC 3'
B 5'UAG 3'
C 5'CUA 3'
D 5'GAU 3'
17,下列哪组密码子是终止密码子
A AAC,ACA,ACU
B AUG,AGU,GAU
C CAA,GAA,UAA
D UAA,UAG,UGA
18,密码子的摆动性表现在
A 反密码子第一位与密码子第三位配对不严格
B 反密码子第三位与密码子第一位配对不严格
C 反密码子第二位与密码子第二位配对不严格
D 反密码子第三位与密码子第三位配对不严格
19,tRNA高级结构为倒L型,其中L长臂的末端是
A 反密码子环
B 氨基酸臂
C 二氢尿嘧啶环
D 附加环
20,真核生物中,参与形成小亚基的rRNA是
A 28S rRNA
B 18S rRNA
C 16S rRNA
D 5S rRNA
21,有关蛋白质合成,下列哪条描述是错误的:
A 基本原料是20种氨基酸
B 直接模板是mRNA
C 合成的方向是从羧基端到氨基端
D 是一个多因子参加的耗能过程
22,下面关于氨酰tRNA合成酶的叙述错误的是
A 催化tRNA3’的羟基与氨基酸羧基形成脂键
B 对氨基酸有高度特异性,对tRNA没有特异性
C 有校正活性
D 活化过程中ATP被水解为AMP
23,下面哪个过程不消耗GTP
A 翻译起始因子释放
B 转肽
C 移位
D 肽链释放
24,翻译起始复合物的组成
A 核糖体+翻译起始因子
B 核糖体+翻译起始因子+mRNA
C 核糖体+mRNA+氨基酰tRNA
D 核糖体+mRNA+甲硫氨酰tRNA
25,翻译起始时与甲酰甲硫氨酰tRNA结合的起始因子是:
A IF1
B IF2
C IF3
D EF-Tu
26,关于移位,错误的是
A 需要EF-G蛋白因子
B 转位完成后,A位空留
C 肽酰tRNA从P位进入A位
D 核糖体延mRNA移动
27,下面关于转肽反应的叙述正确的是
A 肽链从A位tRNA上转移到P位tRNA上
B 需要肽基转移酶的催化
C 转肽反应发生位置在核糖体小亚单位上
D 转肽反应完成后,A位上的tRNA将离开核糖体
28,原核细胞与真核细胞肽链终止因子的数量分别是
A 3,多个
B 1,3
C 3,1
D 1,多个
同课章节目录
- 前言
- 第一章 孟德尔定律
- 第一节 分离定律
- 第二节 自由组合定律
- 第二章 染色体与遗传
- 第一节 减数分裂中的染色体行为
- 第二节 遗传的染色体学说
- 第三节 性染色体与伴性遗传
- 第三章 遗传的分子基础
- 第一节 核酸是遗传物质的证据
- 第二节 DNA的分子结构和特点
- 第三节 遗传信息的传递
- 第四节 遗传信息的表达—-RNA和蛋白质的合成
- 第四章 生物的变异
- 第一节 生物变异的来源
- 第二节 生物变异在生产上的应用
- 第五章 生物的进化
- 第一节 生物的多样性、统一性和进化
- 第二节 进化性变化是怎样发生的
- 第三节 探索生物进化的历史
- 第六章 遗传与人类健康
- 第一节 人类遗传病的主要类型
- 第二节 遗传咨询与优生
- 第三节 基因治疗和人类基因组计划
- 第四节 遗传病与人类未来