癌基因、抑癌基因与生长因子
文档属性
| 名称 | 癌基因、抑癌基因与生长因子 |

|
|
| 格式 | zip | ||
| 文件大小 | 2.7MB | ||
| 资源类型 | 教案 | ||
| 版本资源 | 人教版(新课程标准) | ||
| 科目 | 生物学 | ||
| 更新时间 | 2012-06-02 00:00:00 | ||
图片预览









文档简介
(共45张PPT)
Chapter 20 Oncogene, Anti-oncogene and Growth Factor
第二十章 癌基因、抑癌基因与生长因子
Section 1 Oncogene
第一节 癌基因
癌基因(oncogene)是指存在于正常细胞内,与细胞生长发育调控有关的一组结构基因。
癌基因发生结构异常或表达异常时,可以引起细胞癌变。
癌基因可按其来源不同而分为病毒癌基因(v-onc)和细胞癌基因(c-onc)。
一、病毒癌基因
早在上世纪初(1911年)Reyton Rous 便报道了将鸡肉瘤的无细胞滤液注射给鸡后,可诱导发生肉瘤,说明无细胞滤液中的感染颗粒是引起 Rous鸡肉瘤的原因,这种感染性颗粒后来被证实是逆转录病毒(RNA病毒)。
追踪研究逆转录病毒致癌的线索,Bishop等人于1980年提出了癌基因假说。
根据这一假说,引起Rous鸡肉瘤等的逆转录病毒的基因组中存在有致癌基因,即病毒癌基因,v-onc。
该癌基因来源于正常细胞基因组中的部分序列,即逆转录病毒中的致癌基因序列是在进化过程中通过转导从宿主细胞基因组中获得的。
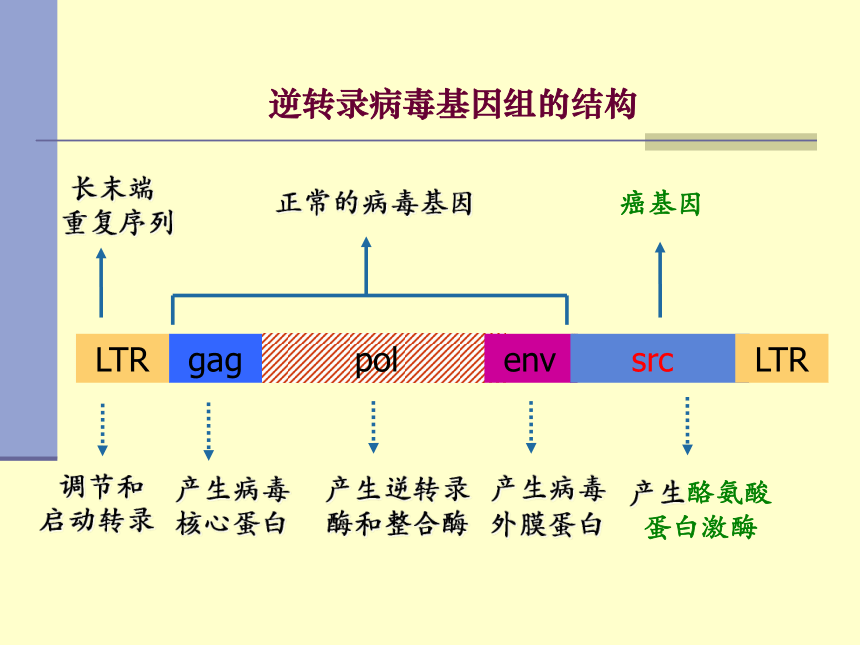
逆转录病毒基因组的结构
调节和
启动转录
LTR
gag
pol
env
src
LTR
长末端
重复序列
癌基因
正常的病毒基因
产生病毒
核心蛋白
产生逆转录
酶和整合酶
产生病毒
外膜蛋白
产生酪氨酸
蛋白激酶
具有致癌性的肿瘤病毒有两种类型:一种是DNA肿瘤病毒,另一种是RNA病毒即逆转录病毒。
DNA肿瘤病毒的基因组的早期功能基因编码转化蛋白,如病毒SV40的 A基因编码大T抗原,分布于胞核,可与p53结合而使之失活。
RNA病毒基因组中可含有致癌基因,并表达相应的转化蛋白,感染动物后即可诱发肿瘤。
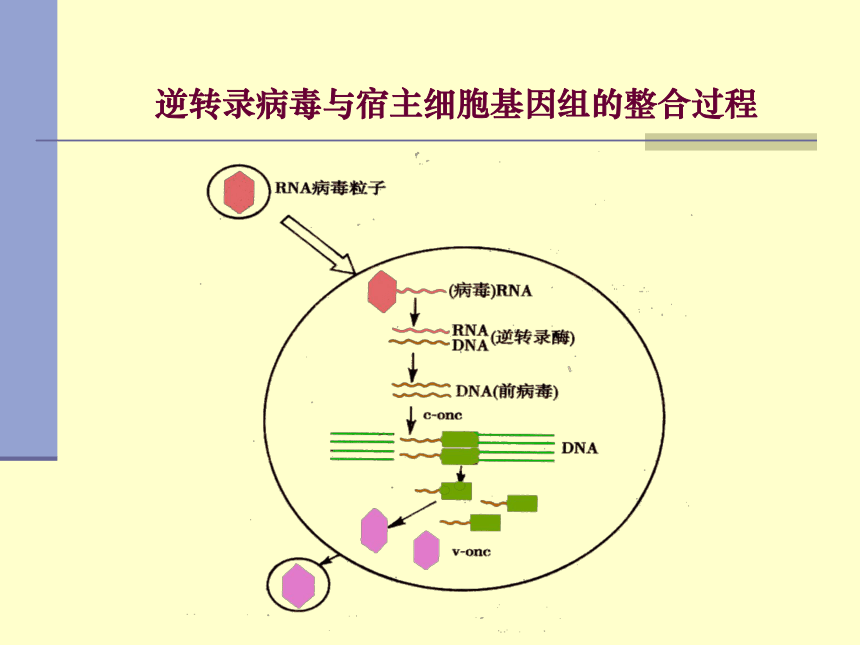
逆转录病毒与宿主细胞基因组的整合过程
因此,病毒癌基因(v-onc)就是指一类存在于肿瘤病毒中,能使宿主细胞发生恶性转化的基因。
二、细胞癌基因
细胞癌基因(c-onc)又称为原癌基因(proto-oncogene),大多数的原癌基因属于调控细胞生长分化的正常基因,原癌基因的蛋白产物是通过影响细胞生长分化中的控制系统而起作用的。
细胞癌基因的特点:
广泛存在于生物界中;
基因序列高度保守;
作用通过其产物蛋白质来体现;
被激活后,形成癌性的细胞转化基因。
目前发现的细胞癌基因已超过100种,根据这些基因表达蛋白产物的功能及亚细胞定位,可将细胞癌基因分为五大类:
1.生长因子类:如c-sis家族癌基因,编码PDGF的 链。
2.信号转导受体类:如erbB癌基因,编码EGF受体;erbA癌基因,编码甲状腺激素受体。
3.G蛋白类:如ras家族,其编码产物为G蛋白,能传递生长信号。
4.信号转导蛋白类:如src家族,其编码产物为细胞内的生长信号传递蛋白,通常含酪氨酸蛋白激酶活性。
5.转录因子类:如myc家族和myb家族,其编码产物为存在于细胞核内的转录因子。
三、癌基因的激活机制
指来源于病毒等的启动子或增强子插入到细胞癌基因的附近或内部而使其开放并异常转录。
如鸡白细胞增生病毒引起的淋巴瘤,就是由于病毒的DNA序列整合到宿主细胞c-myc基因附近,成为该基因的强启动子,导致c-myc基因过强表达。
(一)插入激活
c-myc原癌基因的插入激活
由于染色体重排而导致细胞癌基因从正常位置转移到另一位置,常常是插入一启动子后而使其转录活性增加。
如c-abl原癌基因,经重排后插入到另一称为bcr基因的启动子之后,而使其转录活性增加,从而引起慢性粒细胞性白血病的发生。
(二)基因易位
c-abl原癌基因的易位激活
即基因数量或拷贝数目明显增加。
常见的为myc原癌基因,由于其基因数目的增多而使其表达的蛋白产物增多。
(三)基因扩增
原癌基因的扩增激活
常见的为ras原癌基因的点突变。
ras原癌基因点突变后导致其GTPase活性下降,不能将其结合的GTP迅速水解,从而使其持续保持激活状态。
(四)点突变
四、原癌基因表达产物与生长信息转导
第二节 抑癌基因
Section 2 Anti-oncogene
抑癌基因又称为抗癌基因(anti-oncogene)或肿瘤抑制基因,是指存在于正常细胞中,其编码产物能抑制细胞生长、增殖的一组结构基因。
最初,通过细胞杂交实验,发现并鉴定了抑癌基因。
一、抑癌基因的基本概念
细胞杂交试验
正常细胞
肿瘤细胞
非肿瘤型杂交细胞
杂交融合
肿瘤细胞1
肿瘤细胞2
杂交融合
非肿瘤型杂交细胞
正常细胞
失去某些基因
肿瘤细胞
常见的抑癌基因有Rb基因,p53基因,p16基因等。
其中,Rb基因编码P105Rb转录因子,p53基因编码P53转录因子,p16基因编码一种P16蛋白。
二、常见的抑癌基因
三、抑癌基因的作用机制
Rb基因的失活主要与视网膜母细胞瘤的发生相关。
Rb基因的表达产物为P105Rb,在细胞内存在低磷酸化型(活性型)和高磷酸化型(非活性型)两种类型。
不同类型的P105Rb对转录因子E2F有不同的亲和力。
(一)Rb基因的作用机制
低磷酸化型的P105Rb可与转录因子E2F结合形成复合物,从而阻止E2F启动某些在细胞增殖过程中起主要作用的基因的转录,如c-myc基因等。
高磷酸化型的P105Rb则可促使其与E2F转录因子分离,从而使其呈现活性,细胞即由G1期进入S期。
Rb基因失活后,其表达产物不能与E2F形成复合物,E2F处于持续激活状态,导致细胞增殖。
p105Rb的作用机制(演示)
G0 G1期
Rb蛋白
E-2F
S期
E-2F
DNA
DNA
Rb蛋白
P
mRNA
p53基因的表达产物是分子量为53kD的转录因子P53。
P53蛋白可分为三个结构域:N端的酸性区、中段疏水核心区与C端的碱性区,分子中有一核定位信号,所以它一般都位于核内。
(二)p53基因的作用机制
酸性区
核心区
碱性区
P53蛋白
P53蛋白可与特异的DNA片段结合,其酸性区域具有许多转录因子的共同特征,并与寡聚体的形成有关。
P53蛋白能以四聚体的形式与DNA结合,调节其它基因的表达。
P53蛋白的生物学功能较为复杂,目前已知的功能有:
① 转录调节及抑制细胞生长的作用:
P53蛋白促进WAF1/CIP1基因表达产生一种分子量为21kD的蛋白质(p21WAF1/CIP1),诱导细胞生长停滞于G1期。
② 抑制DNA复制:
P53蛋白可与复制蛋白A(RPA) 结合,而RPA与解链状态的DNA单链的结合可能是DNA复制的起始步骤。
P53蛋白与RPA结合后可抑制RPA与单链DNA的结合,从而抑制了RPA与DNA复制起始点的结合,阻止细胞进入S期。
P53蛋白还可通过抑制解链酶的活性,以抑制DNA复制。
③ 参与程序性细胞凋亡:
P53 蛋白使损伤细胞停留在G1期,以便使DNA损伤修复后再进入细胞周期。若损伤不能修复,则启动细胞进入程序性凋亡(细胞自杀)。
P53蛋白的作用机制(演示)
P53蛋白
解链酶
复制因子A
P21蛋白
细胞停滞于G1期
细胞自杀
DNA损伤
P21基因
P53蛋白
抑 制
P53蛋白
成功修复
修复失败
第三节 生长因子
Section 3 Growth Factor
生长因子(growth factor)是一类通过质膜上特异的受体,将信息传递至细胞内部,调节细胞生长与增殖的多肽或蛋白质类信息物质。
一、概述
生长因子的作用模式 :
内分泌 (endocrine)
旁分泌 (paracrine)
自分泌 (autocrine)
二、生长因子的作用机制
产生相应第二信使
胞内相关蛋白质被磷酸化
与膜受体结合
酪氨酸激酶活化
蛋白激酶活化
核内转录因子活化
基因转录
与胞内受体结合
生长因子 – 受体复合物,活化相关基因
三、生长因子与疾病
细胞凋亡 (apoptosis)在某些生理或病理条下,细胞接受到某种信号所触发的并按一定程序缓慢死亡的过程。
(一)细胞凋亡
诱导因素:野生型p53基因
抑制因素:突变型p53基因
神经生长因子(NGF)
1. 原发性高血压
myc、fos的激活促平滑肌细胞增生;
p53低表达或突变。
2. 动脉粥样硬化
PDGF过量产生。
3. 心肌肥厚
ras、myb、myc、fos等过量表达;
相关的生长因子包括IGF、TGF、FGF等。
(二)心血管疾病
Chapter 20 Oncogene, Anti-oncogene and Growth Factor
第二十章 癌基因、抑癌基因与生长因子
Section 1 Oncogene
第一节 癌基因
癌基因(oncogene)是指存在于正常细胞内,与细胞生长发育调控有关的一组结构基因。
癌基因发生结构异常或表达异常时,可以引起细胞癌变。
癌基因可按其来源不同而分为病毒癌基因(v-onc)和细胞癌基因(c-onc)。
一、病毒癌基因
早在上世纪初(1911年)Reyton Rous 便报道了将鸡肉瘤的无细胞滤液注射给鸡后,可诱导发生肉瘤,说明无细胞滤液中的感染颗粒是引起 Rous鸡肉瘤的原因,这种感染性颗粒后来被证实是逆转录病毒(RNA病毒)。
追踪研究逆转录病毒致癌的线索,Bishop等人于1980年提出了癌基因假说。
根据这一假说,引起Rous鸡肉瘤等的逆转录病毒的基因组中存在有致癌基因,即病毒癌基因,v-onc。
该癌基因来源于正常细胞基因组中的部分序列,即逆转录病毒中的致癌基因序列是在进化过程中通过转导从宿主细胞基因组中获得的。
逆转录病毒基因组的结构
调节和
启动转录
LTR
gag
pol
env
src
LTR
长末端
重复序列
癌基因
正常的病毒基因
产生病毒
核心蛋白
产生逆转录
酶和整合酶
产生病毒
外膜蛋白
产生酪氨酸
蛋白激酶
具有致癌性的肿瘤病毒有两种类型:一种是DNA肿瘤病毒,另一种是RNA病毒即逆转录病毒。
DNA肿瘤病毒的基因组的早期功能基因编码转化蛋白,如病毒SV40的 A基因编码大T抗原,分布于胞核,可与p53结合而使之失活。
RNA病毒基因组中可含有致癌基因,并表达相应的转化蛋白,感染动物后即可诱发肿瘤。
逆转录病毒与宿主细胞基因组的整合过程
因此,病毒癌基因(v-onc)就是指一类存在于肿瘤病毒中,能使宿主细胞发生恶性转化的基因。
二、细胞癌基因
细胞癌基因(c-onc)又称为原癌基因(proto-oncogene),大多数的原癌基因属于调控细胞生长分化的正常基因,原癌基因的蛋白产物是通过影响细胞生长分化中的控制系统而起作用的。
细胞癌基因的特点:
广泛存在于生物界中;
基因序列高度保守;
作用通过其产物蛋白质来体现;
被激活后,形成癌性的细胞转化基因。
目前发现的细胞癌基因已超过100种,根据这些基因表达蛋白产物的功能及亚细胞定位,可将细胞癌基因分为五大类:
1.生长因子类:如c-sis家族癌基因,编码PDGF的 链。
2.信号转导受体类:如erbB癌基因,编码EGF受体;erbA癌基因,编码甲状腺激素受体。
3.G蛋白类:如ras家族,其编码产物为G蛋白,能传递生长信号。
4.信号转导蛋白类:如src家族,其编码产物为细胞内的生长信号传递蛋白,通常含酪氨酸蛋白激酶活性。
5.转录因子类:如myc家族和myb家族,其编码产物为存在于细胞核内的转录因子。
三、癌基因的激活机制
指来源于病毒等的启动子或增强子插入到细胞癌基因的附近或内部而使其开放并异常转录。
如鸡白细胞增生病毒引起的淋巴瘤,就是由于病毒的DNA序列整合到宿主细胞c-myc基因附近,成为该基因的强启动子,导致c-myc基因过强表达。
(一)插入激活
c-myc原癌基因的插入激活
由于染色体重排而导致细胞癌基因从正常位置转移到另一位置,常常是插入一启动子后而使其转录活性增加。
如c-abl原癌基因,经重排后插入到另一称为bcr基因的启动子之后,而使其转录活性增加,从而引起慢性粒细胞性白血病的发生。
(二)基因易位
c-abl原癌基因的易位激活
即基因数量或拷贝数目明显增加。
常见的为myc原癌基因,由于其基因数目的增多而使其表达的蛋白产物增多。
(三)基因扩增
原癌基因的扩增激活
常见的为ras原癌基因的点突变。
ras原癌基因点突变后导致其GTPase活性下降,不能将其结合的GTP迅速水解,从而使其持续保持激活状态。
(四)点突变
四、原癌基因表达产物与生长信息转导
第二节 抑癌基因
Section 2 Anti-oncogene
抑癌基因又称为抗癌基因(anti-oncogene)或肿瘤抑制基因,是指存在于正常细胞中,其编码产物能抑制细胞生长、增殖的一组结构基因。
最初,通过细胞杂交实验,发现并鉴定了抑癌基因。
一、抑癌基因的基本概念
细胞杂交试验
正常细胞
肿瘤细胞
非肿瘤型杂交细胞
杂交融合
肿瘤细胞1
肿瘤细胞2
杂交融合
非肿瘤型杂交细胞
正常细胞
失去某些基因
肿瘤细胞
常见的抑癌基因有Rb基因,p53基因,p16基因等。
其中,Rb基因编码P105Rb转录因子,p53基因编码P53转录因子,p16基因编码一种P16蛋白。
二、常见的抑癌基因
三、抑癌基因的作用机制
Rb基因的失活主要与视网膜母细胞瘤的发生相关。
Rb基因的表达产物为P105Rb,在细胞内存在低磷酸化型(活性型)和高磷酸化型(非活性型)两种类型。
不同类型的P105Rb对转录因子E2F有不同的亲和力。
(一)Rb基因的作用机制
低磷酸化型的P105Rb可与转录因子E2F结合形成复合物,从而阻止E2F启动某些在细胞增殖过程中起主要作用的基因的转录,如c-myc基因等。
高磷酸化型的P105Rb则可促使其与E2F转录因子分离,从而使其呈现活性,细胞即由G1期进入S期。
Rb基因失活后,其表达产物不能与E2F形成复合物,E2F处于持续激活状态,导致细胞增殖。
p105Rb的作用机制(演示)
G0 G1期
Rb蛋白
E-2F
S期
E-2F
DNA
DNA
Rb蛋白
P
mRNA
p53基因的表达产物是分子量为53kD的转录因子P53。
P53蛋白可分为三个结构域:N端的酸性区、中段疏水核心区与C端的碱性区,分子中有一核定位信号,所以它一般都位于核内。
(二)p53基因的作用机制
酸性区
核心区
碱性区
P53蛋白
P53蛋白可与特异的DNA片段结合,其酸性区域具有许多转录因子的共同特征,并与寡聚体的形成有关。
P53蛋白能以四聚体的形式与DNA结合,调节其它基因的表达。
P53蛋白的生物学功能较为复杂,目前已知的功能有:
① 转录调节及抑制细胞生长的作用:
P53蛋白促进WAF1/CIP1基因表达产生一种分子量为21kD的蛋白质(p21WAF1/CIP1),诱导细胞生长停滞于G1期。
② 抑制DNA复制:
P53蛋白可与复制蛋白A(RPA) 结合,而RPA与解链状态的DNA单链的结合可能是DNA复制的起始步骤。
P53蛋白与RPA结合后可抑制RPA与单链DNA的结合,从而抑制了RPA与DNA复制起始点的结合,阻止细胞进入S期。
P53蛋白还可通过抑制解链酶的活性,以抑制DNA复制。
③ 参与程序性细胞凋亡:
P53 蛋白使损伤细胞停留在G1期,以便使DNA损伤修复后再进入细胞周期。若损伤不能修复,则启动细胞进入程序性凋亡(细胞自杀)。
P53蛋白的作用机制(演示)
P53蛋白
解链酶
复制因子A
P21蛋白
细胞停滞于G1期
细胞自杀
DNA损伤
P21基因
P53蛋白
抑 制
P53蛋白
成功修复
修复失败
第三节 生长因子
Section 3 Growth Factor
生长因子(growth factor)是一类通过质膜上特异的受体,将信息传递至细胞内部,调节细胞生长与增殖的多肽或蛋白质类信息物质。
一、概述
生长因子的作用模式 :
内分泌 (endocrine)
旁分泌 (paracrine)
自分泌 (autocrine)
二、生长因子的作用机制
产生相应第二信使
胞内相关蛋白质被磷酸化
与膜受体结合
酪氨酸激酶活化
蛋白激酶活化
核内转录因子活化
基因转录
与胞内受体结合
生长因子 – 受体复合物,活化相关基因
三、生长因子与疾病
细胞凋亡 (apoptosis)在某些生理或病理条下,细胞接受到某种信号所触发的并按一定程序缓慢死亡的过程。
(一)细胞凋亡
诱导因素:野生型p53基因
抑制因素:突变型p53基因
神经生长因子(NGF)
1. 原发性高血压
myc、fos的激活促平滑肌细胞增生;
p53低表达或突变。
2. 动脉粥样硬化
PDGF过量产生。
3. 心肌肥厚
ras、myb、myc、fos等过量表达;
相关的生长因子包括IGF、TGF、FGF等。
(二)心血管疾病
同课章节目录
- 第一章 走近细胞
- 第1节 从生物圈到细胞
- 第2节 细胞的多样性和统一性
- 第二章 组成细胞的分子
- 第1节 细胞中的元素和化合物
- 第2节 生命活动的主要承担者──蛋白质
- 第3节 遗传信息的携带者──核酸
- 第4节 细胞中的糖类和脂质
- 第5节 细胞中的无机物
- 第三章 细胞的基本结构
- 第1节 细胞膜──系统的边界
- 第2节 细胞器──系统内的分工合作
- 第3节 细胞核──系统的控制中心
- 第四章 细胞的物质输入和输出
- 第1节 物质跨膜运输的实例
- 第2节 生物膜的流动镶嵌模型
- 第3节 物质跨膜运输的方式
- 第五章 细胞的能量供应和利用
- 第1节 降低化学反应活化能的酶
- 第2节 细胞的能量“通货”──ATP
- 第3节 ATP的主要来源──细胞呼吸
- 第4节 能量之源——光与光合作用
- 第六章 细胞的生命历程
- 第1节 细胞的增殖
- 第2节 细胞的分化
- 第3节 细胞的衰老和凋亡
- 第4节 细胞的癌变