群落的演替
图片预览









文档简介
(共69张PPT)
群落的演替
(一)演替的概念
演替就是指某一地段上一种生物群落被另一种生物群落所取代的过程。
(二)控制演替的几种主要因素
生物群落的演替是群落内部关系(包括种内和种间关系)与外界环境中各种生态因子综合作用的结果。到目前为止,人们对于演替的机制了解得还不够。要搞清演替过程中每一步发生的原因以及有效地预测演替的方向和速度,还有大量的工作要做。
演体发生部分原因:
1.植物繁殖体的迁移、散布和动物的活动性;
2.群落内部环境的变化;
3.种内与种间关系的改变;
4.外界环境条件的变化;
5.人类的活动。
(三)演替的基本类型
演替类型的划分可以按照不同的原则进行。
1、按照演替发生的时间进程:
(1)世纪演替;(2)长期演替;(3)快速演替。
2、按演替发生的起始条件:
(1)原生演替(primary succession);
(2)次生演替(secondary succession)。
3、按基质的性质:
(1)水生演替系列(hydroseres);
(2)旱生演替系列(xerosres);
(3)中生演替系列(mesoseres) 。
4、按控制演替的主导因素:
(1)内因性演替;(2)外因性演替。
5、按群落代谢特征:可划分为自养性演替和异养性演替 。
布满裂隙的冰川侧碛
自发演替
自发演替的动力来自于生物与它们环境之间的相互作用。原生演替(primary succession)发生在新近形成的基质上,如冰川沉积物。先锋物种的营养物的增加和凋落物的积累为新物种移殖做好了准备。次生演替是由于干扰引起的,如洪水、火灾和人类活动。在这两种自然发生的演替中先锋物种的移殖很快,在较强竞争力的物种入侵之前充分利用空间。耐阴物种(shade-tolerant species)成为阴影处的主宰者,它们是一些较慢的移殖者。
演替—经典模型
生态演替是指在一个自然群落中,物种的组成连续地、单方向地、有顺序地变化。这一顺序被称为是一个演替序列,最后达到的阶段称为顶级。早期的演替阶段,具有先锋物种、低生物量和低营养水平的特征。随着演替的进行,群落的复杂性增加,通常在演替的中期阶段,复杂性达到最大。一个中期的演替群落具有高生物量、高有机营养水平和高的物种多样性。
一年生
杂草
多年生
杂草
灌木
早期演替
树木
晚期演替
树木
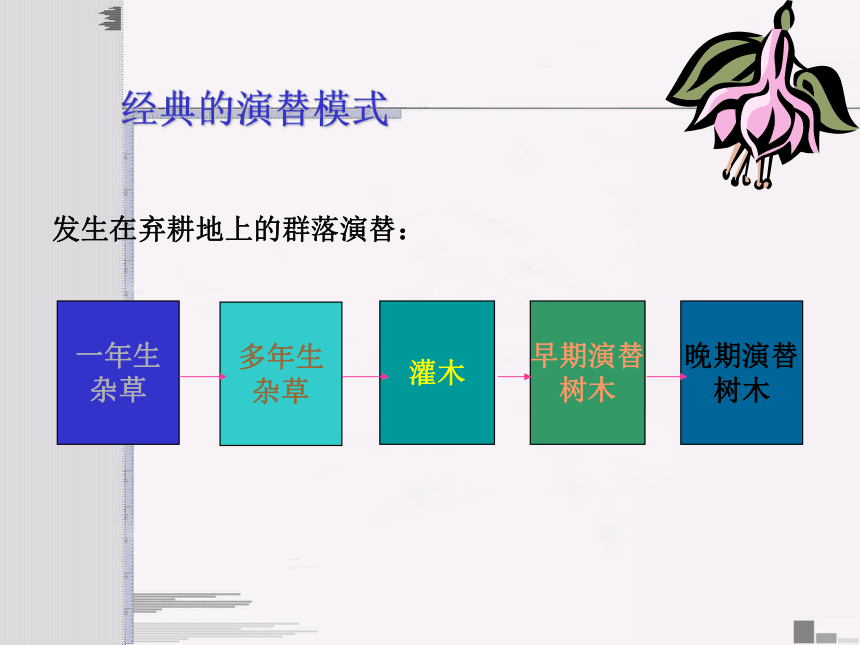
经典的演替模式
发生在弃耕地上的群落演替:
演替序列:如一年生杂草→多年生杂草→灌木→早期演替树木→晚期演替树木,群落的这一顺序被称为一个演替序列,并且每一个明显的和可辨认的阶段都是一个演替系列阶段。
顶级群落:当群落与环境达到稳定平衡状态的时候,演替便放慢了,这个最终的演替系列阶段被称为顶级群落。
先锋物种:早期演替阶段的物种,低生物量和低营养水平。
旱生演替系列:
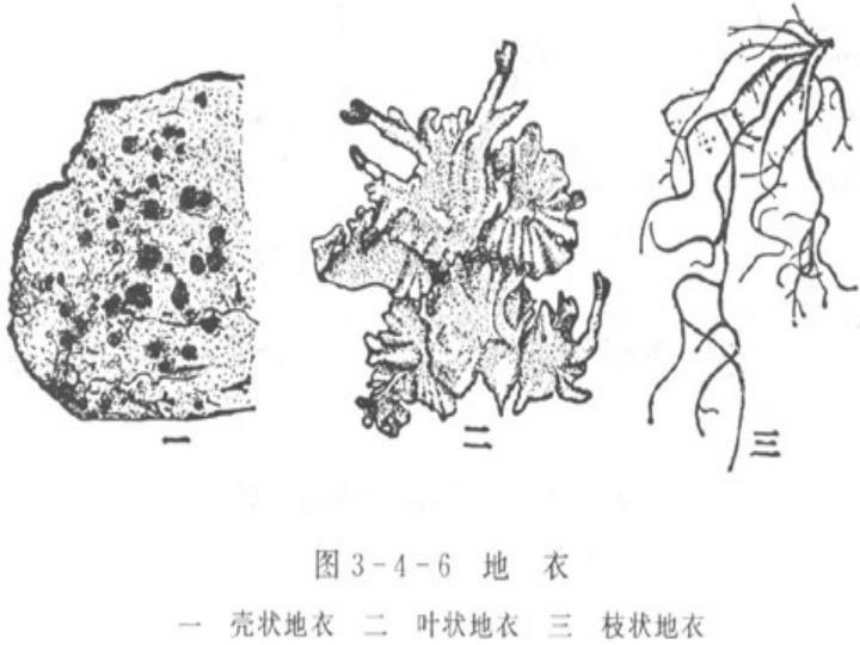
地衣阶段:地衣是在一些裸露的地方首先立足的植物,因为它们能够在极为严酷的自然条件下存活。顺序为:壳状地衣→叶状地衣→枝状地衣。
苔藓阶段:苔藓开始生长在上述的这些浅薄的土壤中,并逐渐取代了先行的植物——地衣,在旱生条件下可以停止生长,进入休眠,应注意的是,各种植物的种子和孢子都会落在这里,并且萌发。问题是在生态选择下,哪些植物可以得到发展。
草本植物阶段:当土壤厚度增加到保持足够湿度的时候,草本植物的幼苗就有了立足点,最后以苔藓取代地衣的同样方式取代了苔藓,使禾草、野菊、紫菀和矮小的木本植物占了优势。
灌木阶段:在这个演替阶段,灌木和小树代替了草本植物。
树木阶段:各种树木在潮湿的、遮荫的地面上生长起来,最终将占有优势。
叶状地衣
壳状地衣
枝状地衣
苔藓阶段
草本阶段
灌木阶段
森林阶段
水生演替系列:
自由漂浮植物阶段 湖底有机质的聚积,主要是依靠浮游有机体的死亡残体,以及湖岸雨水冲刷 使湖底逐渐抬高。
沉水植物阶段 5~7米轮藻属,2~4米金鱼藻、狐尾藻、眼子菜、黑藻、茨藻等。
浮叶根生植物阶段 睡莲科和水鳖科的一些种类,由于这些植物的叶是在水面或水面以上,当它们密集后就将水面完全盖满,使得光照条件变得不利于沉水植物生长,它们就从这里消失而推向水较深的地方。
直立水生植物阶段 主要如芦苇、香蒲、白菖、泽泻等等。其中以芦苇最为常见,其根茎极密,常纠缠绞结,不仅使湖底迅速抬高,而且可以形成一些浮岛。
湿生草本植物阶段 主要是莎草科和禾本科中一些湿生性的种类。在草原地带因为地下水位的降低和地面蒸发的加强,土壤很快变得干燥,湿生草类很快地为旱生草类所代替。
木本植物阶段 首先出现湿性的灌木。而后,随着树木的加入,逐渐形成森林,水位降低,大量地被物也改变了土壤条件,湿生的生境改变成中生生境。
演替实例
动物演替
中国晚中生代以来鱼类区系的演替
美国密执安湖沙丘上的群落演替(原生演替)
裸露沙丘
固沙草本植物(滨草Ammophila breviligulata、沙拂子茅Calamovilfa longifolia)
固沙灌木(沙李Prunus pumila、沙柳Salix spp.、三角杨 Populus deltoides)
松柏林
黑栎林
栎-山核桃林
山毛榉-槭树林
群落演替的实例—湖泊沙丘的群落演替
群落演替的实例—橡果上的异养演替
橡树果提供了生物群落演替的基质
象甲等昆虫侵入橡果,进入橡果胚,在其中产卵,孵化后的幼虫利用橡果胚作为营养;象甲侵入时,亦把真菌带入橡果;
象甲幼虫离开橡果,在果壳上留下洞,食真菌者和食腐动物进入,橡胚组织被降解为粪便;
捕食螨等进入,捕食食腐动物;真菌软化橡果外壳;
较大的动物如毛虫、多足类等进入,橡果崩裂,成土壤腐殖质的一部分。
(五)次生演替
森林的采伐演替
草原的放牧演替也是次生演替中主要的一类。与上述森林采伐演替稍有不同的是:草原放牧演替是逐渐和缓慢发生的。
(一)放牧不足阶段(草原“草甸化”阶段),或完全没有放牧;
(二)轻微放收阶段〔针茅属(Stipa)阶段〕 ;
(三)针茅消灭阶段〔羊茅属(Festuca )阶段〕;
(四)早熟禾废墟阶段 ;
(五)放牧场阶段。
总之,在没有放牧的情况下,草原由于水分条件的改善(因地被物的影响),遭到了中生化作用的结果,亦即草甸化过程;但在强烈放牧情况下,草原遭到了旱生化作用的结果,在草原发育的某些阶段中,草原接近于荒漠带的一些植物群落。这种现象是和水分条件的恶化有关的。土壤在强烈的牲畜践踏下变得坚实,其上层的正常结构遭到破坏,结果土壤的表面蒸发加强,水分情况因此恶化。
十年前的夏尔卓尔
十年后的夏尔卓尔
退行性演替
退行性演替是涉及到移殖和死亡有机体后来腐烂分解的自发演替的一种类型。当有机物质降解耗尽了一些资源和制造了其他可以利用的资源的时候,不同的物种就交替地出现和消失。这个过程导致了腐殖质的生产,并且对土壤形成过程起着重要作用。
进展阶段的特征 逆行阶段的特征
进展阶段和逆行阶段特征的比较
群落结构的复杂化 群落结构的简单化
地面的最大利用 地面的不充分利用
生产力的最大利用 生产力的不充分利用
群落生产率的增加 群落生产率的降低
残遗特有现象的存在,以及对外界环境的适应为方向的物种的形成
新兴特有现象的存在,以及对植物环境的特殊适应为方向的物种形成
群落的中生化 群落的旱生化和湿生化
对外界环境的强烈改造 对外界环境的轻微改造
异发演替
异发演替是由外界环境因素引起的,像长期的气候变化(如冰河时期)和环境在短时间内的变化(如沉积物的增加)。群落结构随时间的变化从沉积物芯样的花粉分析来看是十分明显的。演替进程依赖于物种对环境条件(如盐分)的忍耐性。
演替过程
演替受三个过程的强烈影响: (i)促进:正在发展的群落所产生的非生物环境变化,允许其他物种的入侵。 (ii)抑制:一个阶段的物种抵制后来演替的物种的入侵,以至于入侵仅仅在干扰或死亡后才有可能。
(iii)耐受:后期演替的物种的入侵,因为它仍能够忍受较低的资源水平和排斥早期演替的物种。能忍受低资源水平的强竞争的物种,将取代好的机会迁移者而统治顶极群。
A
B
C
D
促进模型(facilitation model)
物种替代是由于先来物种的活动改变了环境条件,使它不利于自身生存,面促进了后来物种的繁荣;因此物种替代有顺序性,可预测和具方向性。多出现在环境条件严酷的原生演替中。(A、B、C、D代表4个物种,箭头代表被替代)
A
B
C
D
抑制模型(inhibitio model)
先来物种抑制后来物种,使后者难以入侵和发育,因而物种替代没有固定的顺序,各种可能都有,其结果在很大程度上取决于那一种先到。演替在更大程度上决定于个体的生活史对策,因而难以预测。在该模型中没有一个物种可以被认为是竞争的优胜者,而是决定于先到该地,所以演替往往是从短命种到长命种,而不是由规律、可预测的物种替代。(A、B、C、D代表4个物种,箭头代表被替代)
A
B
C
D
忍受模型(tolerance model)
介于上述二者之间,认为物种替代决定于物种的竞争能力。先来的机会种在决定演替途径上并不重要,任何物种都可能开始演替,但有一些物种竞争能力优于其它种,因而它最后能在顶极群落中成为优势种。至于演替的推进是取决于后来入侵还是初始物种的逐渐减少,可能与开始的情形有关。(A、B、C、D代表4个物种,箭头代表被替代)
可定居的空间
只有演替初期
物种能定居
所有生长到成体
的物种能定居
定居的首批物种改变环境
环境不适于早期物种
但较适于后期物种
,早期物种被淘汰
环境较不适于早期物种,
但对于后期物种,既非
较适宜,又非较不适宜
环境不适于所有
物种
最后存留的物种是不
再使环境有利于其它
物种
最后存留的物种能够耐受
早期物种产生的环境变化
,而其它物种无法耐受
最后存留的物种阻
止其它物种侵入,
直到受到干扰为止
干扰破坏顶极阶段
演替开始
促进
耐受
抑制
顶极
顶极
顶极
演替机制
波动和顶级
群落
作为一个地理区域,没有一个单一的顶极群落,但是都有一个连续的顶极类型系列沿着环境梯度变化着。顶极群落是不稳定的,但是它处于一个连续涨落之中。当演替在早期阶段被阻止的时候,朝向顶极的单向演替并不总是发生的。总之,生物量和物种多样性随着演替的进行而增加,但常常在演替中期阶段达到最大,而不是在顶极群落时。
顶极群落的特征物质循环和内稳定性特征
群落特征
物质循环
无机物质循环
生物与环境的物质交换
腐屑在营养物再生中的
作用
内稳定性
内部共生
营养保持
抗干扰能力
熵
信息
演替中群落
顶极群落
开放
快
不重要
不发达
差
弱
高
少
封闭
慢
重要
发达
好
强
低
多
按照Clements的观点,在任何一个地区内,一般的演替系列的终点决定于该地的气候性质,主要表现在顶极群落的优势种,能够很好地适应于地区的气候条件,这样的群落称之为气候顶极群落(Climatic climax),只要是气候保持不急剧的改变,只要没有人类活动和动物的显著影响,或其它侵移方式的发生,它们便一直存在,而且不可能出现任何新的优势植物。这就是所谓的单元顶极(Mono-climax)理论。根据这种理论的解释,一个气候区只有一个潜在的气候顶极群落,这一区域之内的任何一种生境,如给以充分时间,最终都能发展到这种群落。从这个观点出发,势必得出结论,一个气候相当一致的区域,最终将有一种连续的和整齐一致的植被普遍地覆盖着。事实上,在一个气候区域中,总是有土壤的或地形的差异,这些局部环境因素的复合,同普遍的气候环境有很大的差异,在这种生境中,虽然演替进展到稳定的和永久性的群落,但和典型的气候顶极不同,气候顶极可能永远也不会在这种生境中发生。单元顶极的理论没有忽视这些极端的情况,但是比较强调那些预期的情况。
对于各种特殊情况,就分配在另外的类别中。例如特殊的土壤条件(如高度淋溶的灰壤),或者是特殊的地形(如陡坡),或是永久的沼泽等,则发展中的植被可能停止在最后阶段之前的阶段上,也具有较长期的稳定性,则称之为亚顶极(Subclimax)。在某一气候区域中,由于局部环境条件的差异,出现了相邻区域的顶极,则以演替系列中裸地→草本群落→森林的顺序说明其相互关系,而以预顶极(Preclimax前顶极)和超顶极(Post climax后顶极)来加以区别。
例如,在草原区域中,局部出现落叶阔叶林,即为超顶极;而局部出现荒漠类型,则为预顶极。这一现象最明显地表现在山地垂直带上。最后,还有一类情况,即自然顶级被彻底改变为人工群落或天然群落长期用于放牧、割草,它们基本上长期处于人为控制或家畜干扰下,这类情况就被称为干扰顶极(dis-climax)。
Tansley (1954)提出了多元顶极说(Poly-climax theory)。他认为如果一个群落在某种生境中基本稳定,能自行繁殖,并结束了它的演替过程,就可看作是“顶极群落”,而不必汇集于一个共同的气候顶极终点。这样,一个气候区域内可以有多个顶极群落,除了有气候顶极以外,还可以有土壤顶极、地形顶极以及其它等等。因此演替并不导致单一的“气候顶极群落”,而是导致一个顶极群落的镶嵌体,它由相应的生境镶嵌所决定。这就是说,在每一个气候区域内的一个顶极群落(出现在分布最广或“正规的”土壤和地形组合)是气候顶极群落,而在同地区的别的一些群落,则是土壤或地文顶极群落。根据这一概念,任何一个群落,在被任何一个单因素或复合因素稳定到相当长时间的情况下,都可认为是顶极群落。顶级群落之所以能维持不变,是因为它和它的稳定生境之间达到了全部协调的程度。
干 扰
干扰是指林中倒树、食草动物的啃食、潮汐活动、火灾、反常气候变化或人类活动等经常发生的扰乱或干涉,它们迫使物种经历某些选择压力。比如生长在易发生火灾的环境中的植物,在形态和生活史方面有其独特的适应,以保证它们自己和后代在这样的环境中生存下去。
火灾
干扰对物种丰富度的作用
干扰可增加群落的物种多样性,因为它能阻止少数竞争力强的物种成为优势,使其他物种有机会入侵。如果某个地方的种群,由于干扰而不断减少,此地的竞争排斥就可能使那些本来相互竞争的物种停止竞争,而共存在一起。
加拿大一枝黄花具有强干扰作用
中度干扰假说
当干扰为新物种创造了可移殖的斑块的时候,物种多样性趋向最大。但这种情况并不强烈和频繁发生,因而在更大的群落中不会导致物种数的下降。如果干扰小,群落会被一些顶级物种统治。如果干扰很强并经常发生,就会对物理或土壤环境造成影响,从而完全改变群落。
水葫芦
斑块动态
群落的局部性干扰产生了许多断层,相同的或不同的物种个体就乘机移殖到这些断层上来。由于物种移殖到哪个断层是随机的,因此相互竞争的物种可能不会在同一断层上相遇。这样就会使物种丰富的群落生存下来。在晚期演替群落中,如森林群落,断层经历了一个微型演替(mini-succession)的过程,因此形成了一个由处于不同演替阶段的各类斑块组成的镶嵌体。
第五节 植物群落分布规律
植物群落的分布与自然地理环境条件有密切关系,地理环境条件的差异是导致不同植物群落类型及其分布规律的重要原因,不同地区分布的不同植物群落,都是植物群落对该地环境条件的综合反应,是植物群落对该地区环境条件长期适应的历史产物。
垂直地带性,纬度、经度地带性、垂直地带性三者结合,综合影响并决定一个地区的基本特点,或称为“三向地带性学说(三维观点)(three-dimensional view-point)”。
地球上植被的分布,基本上取决于热量与水分这两个因素,并遵循着纬度地带性、经度地带性和垂直地带性而成带地有规律地分布,因此,植被基本上是与整个自然带相吻合。
典型高山植物
一、水平地带性规律
植被的水平地带性是纬度地带性和经度地带性的统称。影响地带性植被的关键因素是热量和水分条件。
我国由于西部有青藏高原,在一定程度上破坏了高空西风环流,中国植物的水平地带性有自身的特点。沿着大兴安岭-吕梁山-六盘山-青藏高原东缘一线,将中国分成东南和西北两半壁,东南半壁是季风区,发育着各类中生性森林,西北部受季风影响小,大多为旱生性的草原和荒漠。在东南半壁,自北向南随着热量的递增,依次出现寒温带针叶林 温带针阔混交林 暖温带落叶阔叶林 亚热带常绿阔叶林(北亚热带落叶常绿阔叶林混交林亚带 中亚热带常绿阔叶林亚带 南亚热带常绿阔叶林亚带) 热带雨林 赤道雨林带。如果沿昆仑山 秦岭 淮河一线以北的温带和暖温带地区,从南到西或从东南到西北,植被依次更替着落叶阔叶林或针阔混交林 草原(草甸草原 典型草原 荒漠草原) 荒漠(草原化荒漠 典型荒漠)。表现了明显的经度地带性。
群落生态学
群落生态学
A.寒温带针叶林区(1.寒温带针叶林地带);
B.温带针阔叶混交林区(2.温带针阔叶混交林地带);
C.暖温带落叶阔叶林区(3.暖温带落叶阔叶林地带);
D.亚热带常绿阔叶林区:D1东部湿润亚区(4.北亚热带常绿落叶阔叶混交林地带,5.中亚热带常绿阔叶林地带,6.南亚热带季风常绿阔叶林地带),D2.西部半湿润亚区(7.中亚热带常绿阔叶林地带,8.南亚热带季风常绿阔叶林地带);
E.热带季雨林、雨林区( 9.热带季雨林、雨林地带);
F.温带草原区:F1.东部亚区,F2.西部亚区;
G.温带荒漠区: G1西部亚区,G2东部亚区;
H.高寒植被区: Hl.山地寒温性针叶林亚区,H2.灌丛、草甸亚区,H3.草原亚区,H4.荒漠亚区
二、垂直地带性
海拔高度不同,气候差异明显。通常海拔高度每上升100m,气温便下降0.5oC左右。在一定范围内,海拔越高,湿度越大,风力越猛,紫外光越强。因此,植被在高山上也表现出与水平面分布的相似性成带现象。垂直地带性是山地植被的重要特点,随着海拔的上升,更替出不同的植被类型。通常一个足够高度的山地,从山麓到山顶更替的植被系列,大致上与从该地所在的水平地带到极地的水平地带的植被系列相似,由山下向山顶分别为森林带、灌木林带、草原带、冻原带,雪线。但是,垂直带与水平带间仅是植被类型间的相似,而非等同。
山地垂直系列中的某些垂直植被带与平地水平系列中的某些水平地带,仍然有着某些种类(特别是科属组成上)、结构特点和生态性质方面或多或少的类似,甚至还有发生上相关性。
我国的山地垂直带谱,大致上可以分为湿润区和干旱区两大类型。
较干旱地区人工植被
湿润地区
群落生态学
长白山的植被垂直带谱:
(1)300~500米,蒙古栎-落叶阔叶杂木林带;
(2)500~1000米,针叶-落叶阔叶混交林带;
(3)1000~1600米,山地针叶林(冷杉林和落叶松林)带;
(4)1600~1900米,山地矮曲林(岳桦林)带;
(5)1900~2744米,山地冻原带。
天山中段北坡的植被垂直带谱:
(1) 500-1000米,荒漠带(包括白琐琐砂荒、假木贼砾质荒漠和蒿属荒漠);
(2)1000-1700米,山地荒漠草原和山地草原带;
(3)1700-2700米,山地针叶林(云杉林)带;
(4)2700-3000米,亚高山草甸带;
(5) 3000-3800米,是高山草甸、高山垫状植被和终年冰雪带。
长白山部分图景
综上所述,可见某处山系上的植披垂直带谱,是反映着该山系所处的一定纬度和一定经度的水平地带的特征,也就是说植被垂直地带性是从属于水平地带性的特征。在水平地带性和垂直地带性的相互关系中,水平地带性是基础,由它决定着山地垂直地带的系统。
群落的演替
(一)演替的概念
演替就是指某一地段上一种生物群落被另一种生物群落所取代的过程。
(二)控制演替的几种主要因素
生物群落的演替是群落内部关系(包括种内和种间关系)与外界环境中各种生态因子综合作用的结果。到目前为止,人们对于演替的机制了解得还不够。要搞清演替过程中每一步发生的原因以及有效地预测演替的方向和速度,还有大量的工作要做。
演体发生部分原因:
1.植物繁殖体的迁移、散布和动物的活动性;
2.群落内部环境的变化;
3.种内与种间关系的改变;
4.外界环境条件的变化;
5.人类的活动。
(三)演替的基本类型
演替类型的划分可以按照不同的原则进行。
1、按照演替发生的时间进程:
(1)世纪演替;(2)长期演替;(3)快速演替。
2、按演替发生的起始条件:
(1)原生演替(primary succession);
(2)次生演替(secondary succession)。
3、按基质的性质:
(1)水生演替系列(hydroseres);
(2)旱生演替系列(xerosres);
(3)中生演替系列(mesoseres) 。
4、按控制演替的主导因素:
(1)内因性演替;(2)外因性演替。
5、按群落代谢特征:可划分为自养性演替和异养性演替 。
布满裂隙的冰川侧碛
自发演替
自发演替的动力来自于生物与它们环境之间的相互作用。原生演替(primary succession)发生在新近形成的基质上,如冰川沉积物。先锋物种的营养物的增加和凋落物的积累为新物种移殖做好了准备。次生演替是由于干扰引起的,如洪水、火灾和人类活动。在这两种自然发生的演替中先锋物种的移殖很快,在较强竞争力的物种入侵之前充分利用空间。耐阴物种(shade-tolerant species)成为阴影处的主宰者,它们是一些较慢的移殖者。
演替—经典模型
生态演替是指在一个自然群落中,物种的组成连续地、单方向地、有顺序地变化。这一顺序被称为是一个演替序列,最后达到的阶段称为顶级。早期的演替阶段,具有先锋物种、低生物量和低营养水平的特征。随着演替的进行,群落的复杂性增加,通常在演替的中期阶段,复杂性达到最大。一个中期的演替群落具有高生物量、高有机营养水平和高的物种多样性。
一年生
杂草
多年生
杂草
灌木
早期演替
树木
晚期演替
树木
经典的演替模式
发生在弃耕地上的群落演替:
演替序列:如一年生杂草→多年生杂草→灌木→早期演替树木→晚期演替树木,群落的这一顺序被称为一个演替序列,并且每一个明显的和可辨认的阶段都是一个演替系列阶段。
顶级群落:当群落与环境达到稳定平衡状态的时候,演替便放慢了,这个最终的演替系列阶段被称为顶级群落。
先锋物种:早期演替阶段的物种,低生物量和低营养水平。
旱生演替系列:
地衣阶段:地衣是在一些裸露的地方首先立足的植物,因为它们能够在极为严酷的自然条件下存活。顺序为:壳状地衣→叶状地衣→枝状地衣。
苔藓阶段:苔藓开始生长在上述的这些浅薄的土壤中,并逐渐取代了先行的植物——地衣,在旱生条件下可以停止生长,进入休眠,应注意的是,各种植物的种子和孢子都会落在这里,并且萌发。问题是在生态选择下,哪些植物可以得到发展。
草本植物阶段:当土壤厚度增加到保持足够湿度的时候,草本植物的幼苗就有了立足点,最后以苔藓取代地衣的同样方式取代了苔藓,使禾草、野菊、紫菀和矮小的木本植物占了优势。
灌木阶段:在这个演替阶段,灌木和小树代替了草本植物。
树木阶段:各种树木在潮湿的、遮荫的地面上生长起来,最终将占有优势。
叶状地衣
壳状地衣
枝状地衣
苔藓阶段
草本阶段
灌木阶段
森林阶段
水生演替系列:
自由漂浮植物阶段 湖底有机质的聚积,主要是依靠浮游有机体的死亡残体,以及湖岸雨水冲刷 使湖底逐渐抬高。
沉水植物阶段 5~7米轮藻属,2~4米金鱼藻、狐尾藻、眼子菜、黑藻、茨藻等。
浮叶根生植物阶段 睡莲科和水鳖科的一些种类,由于这些植物的叶是在水面或水面以上,当它们密集后就将水面完全盖满,使得光照条件变得不利于沉水植物生长,它们就从这里消失而推向水较深的地方。
直立水生植物阶段 主要如芦苇、香蒲、白菖、泽泻等等。其中以芦苇最为常见,其根茎极密,常纠缠绞结,不仅使湖底迅速抬高,而且可以形成一些浮岛。
湿生草本植物阶段 主要是莎草科和禾本科中一些湿生性的种类。在草原地带因为地下水位的降低和地面蒸发的加强,土壤很快变得干燥,湿生草类很快地为旱生草类所代替。
木本植物阶段 首先出现湿性的灌木。而后,随着树木的加入,逐渐形成森林,水位降低,大量地被物也改变了土壤条件,湿生的生境改变成中生生境。
演替实例
动物演替
中国晚中生代以来鱼类区系的演替
美国密执安湖沙丘上的群落演替(原生演替)
裸露沙丘
固沙草本植物(滨草Ammophila breviligulata、沙拂子茅Calamovilfa longifolia)
固沙灌木(沙李Prunus pumila、沙柳Salix spp.、三角杨 Populus deltoides)
松柏林
黑栎林
栎-山核桃林
山毛榉-槭树林
群落演替的实例—湖泊沙丘的群落演替
群落演替的实例—橡果上的异养演替
橡树果提供了生物群落演替的基质
象甲等昆虫侵入橡果,进入橡果胚,在其中产卵,孵化后的幼虫利用橡果胚作为营养;象甲侵入时,亦把真菌带入橡果;
象甲幼虫离开橡果,在果壳上留下洞,食真菌者和食腐动物进入,橡胚组织被降解为粪便;
捕食螨等进入,捕食食腐动物;真菌软化橡果外壳;
较大的动物如毛虫、多足类等进入,橡果崩裂,成土壤腐殖质的一部分。
(五)次生演替
森林的采伐演替
草原的放牧演替也是次生演替中主要的一类。与上述森林采伐演替稍有不同的是:草原放牧演替是逐渐和缓慢发生的。
(一)放牧不足阶段(草原“草甸化”阶段),或完全没有放牧;
(二)轻微放收阶段〔针茅属(Stipa)阶段〕 ;
(三)针茅消灭阶段〔羊茅属(Festuca )阶段〕;
(四)早熟禾废墟阶段 ;
(五)放牧场阶段。
总之,在没有放牧的情况下,草原由于水分条件的改善(因地被物的影响),遭到了中生化作用的结果,亦即草甸化过程;但在强烈放牧情况下,草原遭到了旱生化作用的结果,在草原发育的某些阶段中,草原接近于荒漠带的一些植物群落。这种现象是和水分条件的恶化有关的。土壤在强烈的牲畜践踏下变得坚实,其上层的正常结构遭到破坏,结果土壤的表面蒸发加强,水分情况因此恶化。
十年前的夏尔卓尔
十年后的夏尔卓尔
退行性演替
退行性演替是涉及到移殖和死亡有机体后来腐烂分解的自发演替的一种类型。当有机物质降解耗尽了一些资源和制造了其他可以利用的资源的时候,不同的物种就交替地出现和消失。这个过程导致了腐殖质的生产,并且对土壤形成过程起着重要作用。
进展阶段的特征 逆行阶段的特征
进展阶段和逆行阶段特征的比较
群落结构的复杂化 群落结构的简单化
地面的最大利用 地面的不充分利用
生产力的最大利用 生产力的不充分利用
群落生产率的增加 群落生产率的降低
残遗特有现象的存在,以及对外界环境的适应为方向的物种的形成
新兴特有现象的存在,以及对植物环境的特殊适应为方向的物种形成
群落的中生化 群落的旱生化和湿生化
对外界环境的强烈改造 对外界环境的轻微改造
异发演替
异发演替是由外界环境因素引起的,像长期的气候变化(如冰河时期)和环境在短时间内的变化(如沉积物的增加)。群落结构随时间的变化从沉积物芯样的花粉分析来看是十分明显的。演替进程依赖于物种对环境条件(如盐分)的忍耐性。
演替过程
演替受三个过程的强烈影响: (i)促进:正在发展的群落所产生的非生物环境变化,允许其他物种的入侵。 (ii)抑制:一个阶段的物种抵制后来演替的物种的入侵,以至于入侵仅仅在干扰或死亡后才有可能。
(iii)耐受:后期演替的物种的入侵,因为它仍能够忍受较低的资源水平和排斥早期演替的物种。能忍受低资源水平的强竞争的物种,将取代好的机会迁移者而统治顶极群。
A
B
C
D
促进模型(facilitation model)
物种替代是由于先来物种的活动改变了环境条件,使它不利于自身生存,面促进了后来物种的繁荣;因此物种替代有顺序性,可预测和具方向性。多出现在环境条件严酷的原生演替中。(A、B、C、D代表4个物种,箭头代表被替代)
A
B
C
D
抑制模型(inhibitio model)
先来物种抑制后来物种,使后者难以入侵和发育,因而物种替代没有固定的顺序,各种可能都有,其结果在很大程度上取决于那一种先到。演替在更大程度上决定于个体的生活史对策,因而难以预测。在该模型中没有一个物种可以被认为是竞争的优胜者,而是决定于先到该地,所以演替往往是从短命种到长命种,而不是由规律、可预测的物种替代。(A、B、C、D代表4个物种,箭头代表被替代)
A
B
C
D
忍受模型(tolerance model)
介于上述二者之间,认为物种替代决定于物种的竞争能力。先来的机会种在决定演替途径上并不重要,任何物种都可能开始演替,但有一些物种竞争能力优于其它种,因而它最后能在顶极群落中成为优势种。至于演替的推进是取决于后来入侵还是初始物种的逐渐减少,可能与开始的情形有关。(A、B、C、D代表4个物种,箭头代表被替代)
可定居的空间
只有演替初期
物种能定居
所有生长到成体
的物种能定居
定居的首批物种改变环境
环境不适于早期物种
但较适于后期物种
,早期物种被淘汰
环境较不适于早期物种,
但对于后期物种,既非
较适宜,又非较不适宜
环境不适于所有
物种
最后存留的物种是不
再使环境有利于其它
物种
最后存留的物种能够耐受
早期物种产生的环境变化
,而其它物种无法耐受
最后存留的物种阻
止其它物种侵入,
直到受到干扰为止
干扰破坏顶极阶段
演替开始
促进
耐受
抑制
顶极
顶极
顶极
演替机制
波动和顶级
群落
作为一个地理区域,没有一个单一的顶极群落,但是都有一个连续的顶极类型系列沿着环境梯度变化着。顶极群落是不稳定的,但是它处于一个连续涨落之中。当演替在早期阶段被阻止的时候,朝向顶极的单向演替并不总是发生的。总之,生物量和物种多样性随着演替的进行而增加,但常常在演替中期阶段达到最大,而不是在顶极群落时。
顶极群落的特征物质循环和内稳定性特征
群落特征
物质循环
无机物质循环
生物与环境的物质交换
腐屑在营养物再生中的
作用
内稳定性
内部共生
营养保持
抗干扰能力
熵
信息
演替中群落
顶极群落
开放
快
不重要
不发达
差
弱
高
少
封闭
慢
重要
发达
好
强
低
多
按照Clements的观点,在任何一个地区内,一般的演替系列的终点决定于该地的气候性质,主要表现在顶极群落的优势种,能够很好地适应于地区的气候条件,这样的群落称之为气候顶极群落(Climatic climax),只要是气候保持不急剧的改变,只要没有人类活动和动物的显著影响,或其它侵移方式的发生,它们便一直存在,而且不可能出现任何新的优势植物。这就是所谓的单元顶极(Mono-climax)理论。根据这种理论的解释,一个气候区只有一个潜在的气候顶极群落,这一区域之内的任何一种生境,如给以充分时间,最终都能发展到这种群落。从这个观点出发,势必得出结论,一个气候相当一致的区域,最终将有一种连续的和整齐一致的植被普遍地覆盖着。事实上,在一个气候区域中,总是有土壤的或地形的差异,这些局部环境因素的复合,同普遍的气候环境有很大的差异,在这种生境中,虽然演替进展到稳定的和永久性的群落,但和典型的气候顶极不同,气候顶极可能永远也不会在这种生境中发生。单元顶极的理论没有忽视这些极端的情况,但是比较强调那些预期的情况。
对于各种特殊情况,就分配在另外的类别中。例如特殊的土壤条件(如高度淋溶的灰壤),或者是特殊的地形(如陡坡),或是永久的沼泽等,则发展中的植被可能停止在最后阶段之前的阶段上,也具有较长期的稳定性,则称之为亚顶极(Subclimax)。在某一气候区域中,由于局部环境条件的差异,出现了相邻区域的顶极,则以演替系列中裸地→草本群落→森林的顺序说明其相互关系,而以预顶极(Preclimax前顶极)和超顶极(Post climax后顶极)来加以区别。
例如,在草原区域中,局部出现落叶阔叶林,即为超顶极;而局部出现荒漠类型,则为预顶极。这一现象最明显地表现在山地垂直带上。最后,还有一类情况,即自然顶级被彻底改变为人工群落或天然群落长期用于放牧、割草,它们基本上长期处于人为控制或家畜干扰下,这类情况就被称为干扰顶极(dis-climax)。
Tansley (1954)提出了多元顶极说(Poly-climax theory)。他认为如果一个群落在某种生境中基本稳定,能自行繁殖,并结束了它的演替过程,就可看作是“顶极群落”,而不必汇集于一个共同的气候顶极终点。这样,一个气候区域内可以有多个顶极群落,除了有气候顶极以外,还可以有土壤顶极、地形顶极以及其它等等。因此演替并不导致单一的“气候顶极群落”,而是导致一个顶极群落的镶嵌体,它由相应的生境镶嵌所决定。这就是说,在每一个气候区域内的一个顶极群落(出现在分布最广或“正规的”土壤和地形组合)是气候顶极群落,而在同地区的别的一些群落,则是土壤或地文顶极群落。根据这一概念,任何一个群落,在被任何一个单因素或复合因素稳定到相当长时间的情况下,都可认为是顶极群落。顶级群落之所以能维持不变,是因为它和它的稳定生境之间达到了全部协调的程度。
干 扰
干扰是指林中倒树、食草动物的啃食、潮汐活动、火灾、反常气候变化或人类活动等经常发生的扰乱或干涉,它们迫使物种经历某些选择压力。比如生长在易发生火灾的环境中的植物,在形态和生活史方面有其独特的适应,以保证它们自己和后代在这样的环境中生存下去。
火灾
干扰对物种丰富度的作用
干扰可增加群落的物种多样性,因为它能阻止少数竞争力强的物种成为优势,使其他物种有机会入侵。如果某个地方的种群,由于干扰而不断减少,此地的竞争排斥就可能使那些本来相互竞争的物种停止竞争,而共存在一起。
加拿大一枝黄花具有强干扰作用
中度干扰假说
当干扰为新物种创造了可移殖的斑块的时候,物种多样性趋向最大。但这种情况并不强烈和频繁发生,因而在更大的群落中不会导致物种数的下降。如果干扰小,群落会被一些顶级物种统治。如果干扰很强并经常发生,就会对物理或土壤环境造成影响,从而完全改变群落。
水葫芦
斑块动态
群落的局部性干扰产生了许多断层,相同的或不同的物种个体就乘机移殖到这些断层上来。由于物种移殖到哪个断层是随机的,因此相互竞争的物种可能不会在同一断层上相遇。这样就会使物种丰富的群落生存下来。在晚期演替群落中,如森林群落,断层经历了一个微型演替(mini-succession)的过程,因此形成了一个由处于不同演替阶段的各类斑块组成的镶嵌体。
第五节 植物群落分布规律
植物群落的分布与自然地理环境条件有密切关系,地理环境条件的差异是导致不同植物群落类型及其分布规律的重要原因,不同地区分布的不同植物群落,都是植物群落对该地环境条件的综合反应,是植物群落对该地区环境条件长期适应的历史产物。
垂直地带性,纬度、经度地带性、垂直地带性三者结合,综合影响并决定一个地区的基本特点,或称为“三向地带性学说(三维观点)(three-dimensional view-point)”。
地球上植被的分布,基本上取决于热量与水分这两个因素,并遵循着纬度地带性、经度地带性和垂直地带性而成带地有规律地分布,因此,植被基本上是与整个自然带相吻合。
典型高山植物
一、水平地带性规律
植被的水平地带性是纬度地带性和经度地带性的统称。影响地带性植被的关键因素是热量和水分条件。
我国由于西部有青藏高原,在一定程度上破坏了高空西风环流,中国植物的水平地带性有自身的特点。沿着大兴安岭-吕梁山-六盘山-青藏高原东缘一线,将中国分成东南和西北两半壁,东南半壁是季风区,发育着各类中生性森林,西北部受季风影响小,大多为旱生性的草原和荒漠。在东南半壁,自北向南随着热量的递增,依次出现寒温带针叶林 温带针阔混交林 暖温带落叶阔叶林 亚热带常绿阔叶林(北亚热带落叶常绿阔叶林混交林亚带 中亚热带常绿阔叶林亚带 南亚热带常绿阔叶林亚带) 热带雨林 赤道雨林带。如果沿昆仑山 秦岭 淮河一线以北的温带和暖温带地区,从南到西或从东南到西北,植被依次更替着落叶阔叶林或针阔混交林 草原(草甸草原 典型草原 荒漠草原) 荒漠(草原化荒漠 典型荒漠)。表现了明显的经度地带性。
群落生态学
群落生态学
A.寒温带针叶林区(1.寒温带针叶林地带);
B.温带针阔叶混交林区(2.温带针阔叶混交林地带);
C.暖温带落叶阔叶林区(3.暖温带落叶阔叶林地带);
D.亚热带常绿阔叶林区:D1东部湿润亚区(4.北亚热带常绿落叶阔叶混交林地带,5.中亚热带常绿阔叶林地带,6.南亚热带季风常绿阔叶林地带),D2.西部半湿润亚区(7.中亚热带常绿阔叶林地带,8.南亚热带季风常绿阔叶林地带);
E.热带季雨林、雨林区( 9.热带季雨林、雨林地带);
F.温带草原区:F1.东部亚区,F2.西部亚区;
G.温带荒漠区: G1西部亚区,G2东部亚区;
H.高寒植被区: Hl.山地寒温性针叶林亚区,H2.灌丛、草甸亚区,H3.草原亚区,H4.荒漠亚区
二、垂直地带性
海拔高度不同,气候差异明显。通常海拔高度每上升100m,气温便下降0.5oC左右。在一定范围内,海拔越高,湿度越大,风力越猛,紫外光越强。因此,植被在高山上也表现出与水平面分布的相似性成带现象。垂直地带性是山地植被的重要特点,随着海拔的上升,更替出不同的植被类型。通常一个足够高度的山地,从山麓到山顶更替的植被系列,大致上与从该地所在的水平地带到极地的水平地带的植被系列相似,由山下向山顶分别为森林带、灌木林带、草原带、冻原带,雪线。但是,垂直带与水平带间仅是植被类型间的相似,而非等同。
山地垂直系列中的某些垂直植被带与平地水平系列中的某些水平地带,仍然有着某些种类(特别是科属组成上)、结构特点和生态性质方面或多或少的类似,甚至还有发生上相关性。
我国的山地垂直带谱,大致上可以分为湿润区和干旱区两大类型。
较干旱地区人工植被
湿润地区
群落生态学
长白山的植被垂直带谱:
(1)300~500米,蒙古栎-落叶阔叶杂木林带;
(2)500~1000米,针叶-落叶阔叶混交林带;
(3)1000~1600米,山地针叶林(冷杉林和落叶松林)带;
(4)1600~1900米,山地矮曲林(岳桦林)带;
(5)1900~2744米,山地冻原带。
天山中段北坡的植被垂直带谱:
(1) 500-1000米,荒漠带(包括白琐琐砂荒、假木贼砾质荒漠和蒿属荒漠);
(2)1000-1700米,山地荒漠草原和山地草原带;
(3)1700-2700米,山地针叶林(云杉林)带;
(4)2700-3000米,亚高山草甸带;
(5) 3000-3800米,是高山草甸、高山垫状植被和终年冰雪带。
长白山部分图景
综上所述,可见某处山系上的植披垂直带谱,是反映着该山系所处的一定纬度和一定经度的水平地带的特征,也就是说植被垂直地带性是从属于水平地带性的特征。在水平地带性和垂直地带性的相互关系中,水平地带性是基础,由它决定着山地垂直地带的系统。
同课章节目录
- 第1章 人体的内环境与稳态
- 第1节 细胞生活的环境
- 第2节 内环境稳态的重要性
- 第2章 动物和人体生命活动的调节
- 第1节 通过神经系统的调节
- 第2节 通过激素的调节
- 第3节 神经调节与体液调节的关系
- 第4节 免疫调节
- 第3章 植物的激素调节
- 第1节 植物生长素的发现
- 第2节 生长素的生理作用
- 第3节 其他植物激素
- 第4章 种群和群落
- 第1节 种群的特征
- 第2节 种群数量的变化
- 第3节 群落的结构
- 第4节 群落的演替
- 第5章 生态系统及其稳定性
- 第1节 生态系统的结构
- 第2节 生态系统的能量流动
- 第3节 生态系统的物质循环
- 第4节 生态系统的信息传递
- 第5节 生态系统的稳定性
- 第6章 生态环境的保护
- 第1节 人口增长对生态环境的影响
- 第2节 保护我们共同的家园